You might also like
- Development of Human LocomotionDocument7 pagesDevelopment of Human LocomotionNaveen KumarNo ratings yet
- Distinct Locomotor Precursors in Newborn BabiesDocument9 pagesDistinct Locomotor Precursors in Newborn BabiesNaveen KumarNo ratings yet
- Development of Human LocomotionDocument7 pagesDevelopment of Human LocomotionNaveen KumarNo ratings yet
- Discovering Research Priorities - Appendices-CompleteDocument16 pagesDiscovering Research Priorities - Appendices-CompleteNaveen KumarNo ratings yet
- Saudi SurveyDocument12 pagesSaudi SurveyNaveen KumarNo ratings yet
- Itr-1 Sahaj Indian Income Tax Return: Acknowledgement Number: Assessment Year: 2020-21Document7 pagesItr-1 Sahaj Indian Income Tax Return: Acknowledgement Number: Assessment Year: 2020-21Prérńã K K ŚhãhNo ratings yet
- Dry Needling As A Treatment Modality For Tendinopathy: A Narrative ReviewDocument8 pagesDry Needling As A Treatment Modality For Tendinopathy: A Narrative ReviewNaveen KumarNo ratings yet
- Saudi SurveyDocument12 pagesSaudi SurveyNaveen KumarNo ratings yet
- Effectiveness of Physical Therapy in Patients With Tension-Type Headache: Literature ReviewDocument8 pagesEffectiveness of Physical Therapy in Patients With Tension-Type Headache: Literature Reviewratnayanti udinNo ratings yet
- Burn Out SyndromeDocument7 pagesBurn Out SyndromeNaveen KumarNo ratings yet
- 373-Article Text-714-1-10-20190608Document4 pages373-Article Text-714-1-10-20190608Naveen KumarNo ratings yet
- Goni OmeterDocument57 pagesGoni OmeterNaveen KumarNo ratings yet
- Factors Affecting The Extent of Utilization of PTDocument7 pagesFactors Affecting The Extent of Utilization of PTNaveen KumarNo ratings yet
- Cochrane: LibraryDocument47 pagesCochrane: LibraryNaveen KumarNo ratings yet
- PEDro ScaleDocument2 pagesPEDro Scalesweetpudding0% (1)
- BERG BALANCE TESTS AND RATING SCALEDocument4 pagesBERG BALANCE TESTS AND RATING SCALEINDO' NURULNo ratings yet
- Dry Needling As A Treatment Modality For Tendinopathy: A Narrative ReviewDocument8 pagesDry Needling As A Treatment Modality For Tendinopathy: A Narrative ReviewNaveen KumarNo ratings yet
- Multiple Sclerosis Impact Scale (MSIS-29)Document2 pagesMultiple Sclerosis Impact Scale (MSIS-29)Naveen Kumar100% (1)
- The Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Document4 pagesThe Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Naveen KumarNo ratings yet
- Functional Independence Measure (FIM ) InstrumentDocument2 pagesFunctional Independence Measure (FIM ) InstrumentAspida100% (4)
- Sy Ringo My EliaDocument14 pagesSy Ringo My EliaNaveen KumarNo ratings yet
- The Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Document4 pagesThe Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Naveen KumarNo ratings yet
- Dynamic Gait IndexDocument3 pagesDynamic Gait IndexÂdâm JônêsNo ratings yet
- FIM ManualDocument24 pagesFIM ManualAnshuman MihirNo ratings yet
- Sy Ringo My EliaDocument14 pagesSy Ringo My EliaNaveen KumarNo ratings yet
- Goni OmeterDocument57 pagesGoni OmeterNaveen KumarNo ratings yet
- Physiotherapy in Pre and Post Operative Spine SurgeriesDocument45 pagesPhysiotherapy in Pre and Post Operative Spine SurgeriesNaveen KumarNo ratings yet
- Developmental DelayDocument79 pagesDevelopmental DelayNaveen KumarNo ratings yet
- Physiotherapy in Pre and Post Operative Spine SurgeriesDocument45 pagesPhysiotherapy in Pre and Post Operative Spine SurgeriesNaveen KumarNo ratings yet
- The Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Document4 pagesThe Amyotrophic Lateral Sclerosis Functional Rating Scale (ALSFRS)Naveen KumarNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Correction Traumatic LPDocument2 pagesCorrection Traumatic LPFitrisia 'pipit' AmelinNo ratings yet
- Craniosacral OsteopathyDocument90 pagesCraniosacral Osteopathykhorrami4100% (6)
- 3 Brain Spinal Cord and Nerve Disorders of DogsDocument18 pages3 Brain Spinal Cord and Nerve Disorders of DogsKoleen Lopez ÜNo ratings yet
- Intersections: Brain Research Foundation 2018-2019 Impact ReportDocument21 pagesIntersections: Brain Research Foundation 2018-2019 Impact ReportDima BoichenkoNo ratings yet
- What Is Nervous System?: Presented By: TIP Grade 12 StudentDocument28 pagesWhat Is Nervous System?: Presented By: TIP Grade 12 StudentDreiza Patria SunodanNo ratings yet
- S.C.E.N.A.R.: Self-Controlled Energetic Neuro-Adaptive RegulatorDocument41 pagesS.C.E.N.A.R.: Self-Controlled Energetic Neuro-Adaptive RegulatorHoracio CastroNo ratings yet
- Lecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesDocument119 pagesLecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without Notesellie marcusNo ratings yet
- 5090 w16 QP CompleteDocument120 pages5090 w16 QP CompleteAyra MujibNo ratings yet
- Nervous System Anatomy and Physiology LectureDocument105 pagesNervous System Anatomy and Physiology LectureButch Dumdum75% (12)
- Perdev Week 5Document3 pagesPerdev Week 5AdiraNo ratings yet
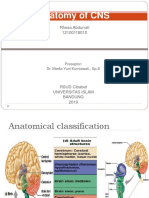
- Anatomy of CNS: Rhesa Abdurrafi 12100118010Document69 pagesAnatomy of CNS: Rhesa Abdurrafi 12100118010Rhessa AbdurrafiNo ratings yet
- محلول mcq's من المريخ فريش NeurophysiologyDocument39 pagesمحلول mcq's من المريخ فريش Neurophysiologyleen100% (1)
- Ericsson 2G&3G Engineering Parameter-15rd DEC V12Document616 pagesEricsson 2G&3G Engineering Parameter-15rd DEC V12Muhammad Ikhsan Paripurna50% (2)
- Differences Between Men and Women (Brain)Document21 pagesDifferences Between Men and Women (Brain)CLAIRE DENISSE DEVISNo ratings yet
- Skripta Za Usmeni Iz EngleskogDocument15 pagesSkripta Za Usmeni Iz EngleskogElizabetaNo ratings yet
- Anatomy and Physiology (OpenStax College)Document1,313 pagesAnatomy and Physiology (OpenStax College)Noa Goyanes100% (1)
- (ANA) 5.06 Organization of The Nervous SystemDocument19 pages(ANA) 5.06 Organization of The Nervous SystemAdrian MaterumNo ratings yet
- SCIENCEDocument3 pagesSCIENCENel BorniaNo ratings yet
- Nervous System WorksheetDocument2 pagesNervous System WorksheetHAA-BEENo ratings yet
- Guide to Neuropathology SubspecialtyDocument3 pagesGuide to Neuropathology SubspecialtyYağışBeyNo ratings yet
- Earth and Life Science - Q2 - Mod13 - Characteristics of Organ Systems - Version1Document28 pagesEarth and Life Science - Q2 - Mod13 - Characteristics of Organ Systems - Version1Glenda Astodillo100% (1)
- Chapter 8, Part B - The Spinal Cord!Document9 pagesChapter 8, Part B - The Spinal Cord!Putamen NongchaiNo ratings yet
- Physiology of Nerve Fibers: Dr. Zara BatoolDocument30 pagesPhysiology of Nerve Fibers: Dr. Zara BatoolZuhaib AhmedNo ratings yet
- BiofeedbackDocument105 pagesBiofeedbackspeedy776780% (5)
- Jacaranda 1 - 2 7th Ed PDFDocument496 pagesJacaranda 1 - 2 7th Ed PDFPatricia WillowNo ratings yet
- Brain Anatomy and How The Brain WorksDocument1 pageBrain Anatomy and How The Brain Worksdudong borjaNo ratings yet
- Boron - Medical PhysiologyDocument13 pagesBoron - Medical PhysiologyOlgaBulat0% (6)
- Daftar PustakaDocument3 pagesDaftar PustakaEdwin Batara SaragihNo ratings yet
- The Nervous System: Elaine N. MariebDocument92 pagesThe Nervous System: Elaine N. MariebhamidiNo ratings yet
- ISAP Lecture 1 Module 1 - Studocu NotesDocument6 pagesISAP Lecture 1 Module 1 - Studocu NotesLyndonNo ratings yet