You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- 2023 02 11NewScientistDocument60 pages2023 02 11NewScientistSalahuddin NizamaniNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
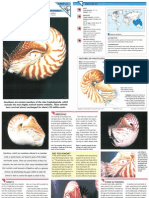
- The Magick of SeashellsDocument6 pagesThe Magick of SeashellsBarbra AntonNo ratings yet
- (1901) Tales of A Dying Race (Maori People of New Zealand)Document300 pages(1901) Tales of A Dying Race (Maori People of New Zealand)Herbert Hillary Booker 2nd100% (4)
- Cephalopods of The World Vol.1 Chambered Nautiluses and SepioidsDocument294 pagesCephalopods of The World Vol.1 Chambered Nautiluses and Sepioidscavris100% (1)
- FAO SPECIES CATALOGUE. Cephalopods of The WorldDocument290 pagesFAO SPECIES CATALOGUE. Cephalopods of The WorldDavid MasallesNo ratings yet
- Wildlife Fact File - Primitive Animals - Pgs. 31-40Document20 pagesWildlife Fact File - Primitive Animals - Pgs. 31-40ClearMind8450% (4)
- AmmonitesDocument156 pagesAmmonitesFlorin Ivan100% (3)
- EntieaelfuiDocument22 pagesEntieaelfuiKeiber GonzalezNo ratings yet
- Canfialidad en SurDocument17 pagesCanfialidad en SurKeiber GonzalezNo ratings yet
- Antigua Grecia FFFDocument7 pagesAntigua Grecia FFFKeiber GonzalezNo ratings yet
- Quimico DelatadDocument10 pagesQuimico DelatadKeiber GonzalezNo ratings yet
- Polancos ReaBeth Video ReviewDocument2 pagesPolancos ReaBeth Video ReviewRea PolancosNo ratings yet
- Skytron Nautilus LEDDocument4 pagesSkytron Nautilus LEDsigmakarsaNo ratings yet
- Inky The Octopus Based On A Real Life Aquatic Escape Erin Guendelsberger David Leonard Full ChapterDocument58 pagesInky The Octopus Based On A Real Life Aquatic Escape Erin Guendelsberger David Leonard Full Chapterkay.cheung868100% (7)
- Amore, Khan - The Divine ProportionDocument57 pagesAmore, Khan - The Divine Proportionpic2007No ratings yet
- Afaq AssignmentDocument9 pagesAfaq AssignmentAhsan Bin KhalidNo ratings yet
- Cambridge IGCSE: Biology 0610/62Document12 pagesCambridge IGCSE: Biology 0610/62Arqam Javed AhmadNo ratings yet
- Bivalvia & Cephalopoda: WelcomeDocument9 pagesBivalvia & Cephalopoda: WelcomeJohn Michael AnselmoNo ratings yet
- LAU Paleoart Workbook - 2023Document16 pagesLAU Paleoart Workbook - 2023samuelaguilar990No ratings yet
- MolluscaDocument35 pagesMolluscaBio SciencesNo ratings yet
- NautilusDocument15 pagesNautilusenzo abrahamNo ratings yet
- 10 Things in Nature That Contains MathDocument1 page10 Things in Nature That Contains MathKaye PauleNo ratings yet
- Geology and PaleontologyDocument184 pagesGeology and PaleontologyGonzalo Galvez CardenasNo ratings yet
- Cephalopod - WikipediaDocument172 pagesCephalopod - WikipediaReynaLord Rafael-sanNo ratings yet
- The Molluscan Bauplan PDFDocument14 pagesThe Molluscan Bauplan PDFNurul MagfirahNo ratings yet
- Class Cephalopods: - Squids, - Cuttle Fish, - Nautilus, - Octopus, EtcDocument18 pagesClass Cephalopods: - Squids, - Cuttle Fish, - Nautilus, - Octopus, EtcAbere TesfayeNo ratings yet
- Nautilus PompiliusDocument6 pagesNautilus PompiliusNicolas DemidovNo ratings yet
- Cephalopod Classification and Taxonomy: V. Venkatesan and K.S. MohamedDocument5 pagesCephalopod Classification and Taxonomy: V. Venkatesan and K.S. MohamedGET ISLAMIC VIDEOSNo ratings yet
- CollectionDocument133 pagesCollectionvinorexNo ratings yet
- Identification of The Middle Eocene Nautiloid Genera of Qatar, Middle-EastDocument327 pagesIdentification of The Middle Eocene Nautiloid Genera of Qatar, Middle-EastJacques LeBlancNo ratings yet
- Phylum MolluscaDocument50 pagesPhylum MolluscaIsaacNo ratings yet