You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Section A.: Basement MembraneDocument6 pagesSection A.: Basement MembraneChong AiklongNo ratings yet
- Dermatologic Immunity PDFDocument391 pagesDermatologic Immunity PDFChino UnjbgNo ratings yet
- The Epidemiological Characteristic and Trends of Burns GloballyDocument16 pagesThe Epidemiological Characteristic and Trends of Burns GloballyFelp ScholzNo ratings yet
- Campbell 2000Document6 pagesCampbell 2000Felp ScholzNo ratings yet
- ThetramfDocument80 pagesThetramfFelp ScholzNo ratings yet
- Wound Healing Activities of Different Extracts ofDocument8 pagesWound Healing Activities of Different Extracts ofFelp ScholzNo ratings yet
- Pab MS 14713 Dec4Document9 pagesPab MS 14713 Dec4Felp ScholzNo ratings yet
- A Systematic Review of The Effect of Centella Asiatica On Wound HealingDocument14 pagesA Systematic Review of The Effect of Centella Asiatica On Wound HealingFelp ScholzNo ratings yet
- Moisturizing Effect of Topical Cosmetic Products Applied To Dry Skin PDFDocument12 pagesMoisturizing Effect of Topical Cosmetic Products Applied To Dry Skin PDFFelp ScholzNo ratings yet
- A Questionnaire For The Assessment of Pruritus: Validation in Uremic PatientsDocument4 pagesA Questionnaire For The Assessment of Pruritus: Validation in Uremic PatientsFelp ScholzNo ratings yet
- Gyanendra TripathiDocument6 pagesGyanendra TripathiFelp ScholzNo ratings yet
- Arnica Montana L. - A Plant of Healing: Review: April 2017Document22 pagesArnica Montana L. - A Plant of Healing: Review: April 2017Felp ScholzNo ratings yet
- Antioxidants 06 00002 v2Document16 pagesAntioxidants 06 00002 v2Felp ScholzNo ratings yet
- Pittler Et Al-2012-Cochrane Database of Systematic ReviewsDocument3 pagesPittler Et Al-2012-Cochrane Database of Systematic ReviewsFelp ScholzNo ratings yet
- Equisetum Arvense (Common Horsetail) Modulates The Function of Inflammatory Immunocompetent CellsDocument10 pagesEquisetum Arvense (Common Horsetail) Modulates The Function of Inflammatory Immunocompetent CellsFelp ScholzNo ratings yet
- Moisturizing Effect of Topical Cosmetic Products Applied To Dry SkinDocument12 pagesMoisturizing Effect of Topical Cosmetic Products Applied To Dry SkinFelp ScholzNo ratings yet
- INDONESIAN JOURNAL OF CLINICAL PATHOLOGY AND MEDICAL LABORATORY Majalah Patologi Klinik Indonesia Dan Laboratorium MedikDocument11 pagesINDONESIAN JOURNAL OF CLINICAL PATHOLOGY AND MEDICAL LABORATORY Majalah Patologi Klinik Indonesia Dan Laboratorium MedikAkputra BinsistNo ratings yet
- Gibco Poly D Lysine FlyerDocument4 pagesGibco Poly D Lysine FlyerW DongNo ratings yet
- Accepted Manuscript: Environmental and Experimental BotanyDocument29 pagesAccepted Manuscript: Environmental and Experimental BotanyRenganathan PrabhaharanNo ratings yet
- ds45 Intro To Cellular OrganellesDocument10 pagesds45 Intro To Cellular Organellesapi-110789702No ratings yet
- WEEK3 ModuleLec (BIO101)Document10 pagesWEEK3 ModuleLec (BIO101)Maricris GatdulaNo ratings yet
- INTERFEROMICSDocument15 pagesINTERFEROMICSAnshul Joshi100% (1)
- Overview of Hemostasis and ThrombosisDocument12 pagesOverview of Hemostasis and ThrombosisMichaella AlmirahNo ratings yet
- Xenopus Laevis and Treating Them With Progesterone. When Cytoplasm From Eggs Arrested in Metaphase ofDocument6 pagesXenopus Laevis and Treating Them With Progesterone. When Cytoplasm From Eggs Arrested in Metaphase ofstevensb055No ratings yet
- Bio Exam Question New OfficialDocument3 pagesBio Exam Question New OfficialnabilahNo ratings yet
- Cell Membranes & Transport 1Document77 pagesCell Membranes & Transport 1Alen SheenNo ratings yet
- Histology Sample ExamsDocument6 pagesHistology Sample ExamsBaluya MohanNo ratings yet
- Dr. Lamia El Wakeel, PhD. Lecturer of Clinical Pharmacy Ain Shams UniversityDocument19 pagesDr. Lamia El Wakeel, PhD. Lecturer of Clinical Pharmacy Ain Shams UniversitysamvetNo ratings yet
- KMPHDocument14 pagesKMPHHaifa amirahNo ratings yet
- ATP Photosynthesis and Cell Respiration Web QuestDocument3 pagesATP Photosynthesis and Cell Respiration Web QuestJakie UbinaNo ratings yet
- The Influence of Anti-Inflammatory Medication On Exercise-Induced Myogenic Precursor Cell Responses in HumansDocument8 pagesThe Influence of Anti-Inflammatory Medication On Exercise-Induced Myogenic Precursor Cell Responses in Humanssaulo_martorelliNo ratings yet
- 2.7 Dna Replication Transcription and Translation 4Document168 pages2.7 Dna Replication Transcription and Translation 4Senam DzakpasuNo ratings yet
- Hemostasis - TrombositDocument28 pagesHemostasis - TrombositNovi AndrianiNo ratings yet
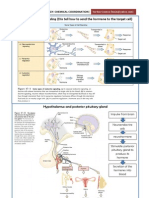
- The Chemical Coordination SummaryDocument7 pagesThe Chemical Coordination SummaryTee Bio-ScienceNo ratings yet
- Atp SynthesisDocument5 pagesAtp Synthesiskman0722No ratings yet
- Zoology Reviewer 1Document3 pagesZoology Reviewer 1Jojit Espiritu100% (1)
- Reviewing Mitosis Discussion QuestionsDocument3 pagesReviewing Mitosis Discussion Questionsapi-521773978No ratings yet
- Biology Midterm Review 2023Document8 pagesBiology Midterm Review 2023beegirl954No ratings yet
- tmpDC1A TMPDocument43 pagestmpDC1A TMPFrontiersNo ratings yet
- Grace's Insect MediumDocument1 pageGrace's Insect MediumAlleleBiotechNo ratings yet
- Ethylene Plant Hormone PDFDocument2 pagesEthylene Plant Hormone PDFLorriNo ratings yet
- Cells AssignmentDocument18 pagesCells AssignmentMuqadasNo ratings yet
- Organization Replication Repair: DR S K BansalDocument71 pagesOrganization Replication Repair: DR S K BansalSanjiv BansalNo ratings yet
- Fatty Acid Synthesis by Prof DR Abdalla Jarari 2nd Year For VIDEOSDocument66 pagesFatty Acid Synthesis by Prof DR Abdalla Jarari 2nd Year For VIDEOSnoran alfaitoryNo ratings yet