You might also like
- Helminth and Diabetes2012Document15 pagesHelminth and Diabetes2012esraaNo ratings yet
- A Teleomorph Is: 3. Which of The Following Statements Best Describe Fungi?Document2 pagesA Teleomorph Is: 3. Which of The Following Statements Best Describe Fungi?esraaNo ratings yet
- Mycology Exam One 8/4/2020 11:30-12:10Document3 pagesMycology Exam One 8/4/2020 11:30-12:10esraaNo ratings yet
- ASTMHExam Brochure 16 FNLV 2Document17 pagesASTMHExam Brochure 16 FNLV 2esraaNo ratings yet
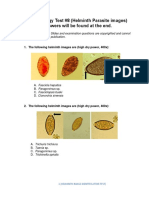
- Parasitology Test #8 (Helminth Parasite Images) Answers Will Be Found at The EndDocument14 pagesParasitology Test #8 (Helminth Parasite Images) Answers Will Be Found at The EndesraaNo ratings yet
- Parasitology Study GuideDocument16 pagesParasitology Study GuideesraaNo ratings yet
- ExamDocument12 pagesExamesraaNo ratings yet
- For ExamDocument9 pagesFor ExamesraaNo ratings yet
- QestionsDocument12 pagesQestionsesraaNo ratings yet
- QestionsDocument12 pagesQestionsesraaNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- (BHU) DoneDocument6 pages(BHU) Donedr.venkat r singhNo ratings yet
- Ouabain - The Optimal Solution For The Problem of Myocardial InfarctionDocument20 pagesOuabain - The Optimal Solution For The Problem of Myocardial InfarctionredsbooksNo ratings yet
- HS Instructor ManualDocument122 pagesHS Instructor ManualJames Cojab SacalNo ratings yet
- Triage Assessment of Psychiatric Patient LectureDocument31 pagesTriage Assessment of Psychiatric Patient Lectureهشام عفانه100% (1)
- Dog Bite Show NotesDocument5 pagesDog Bite Show NotesLakshay ChananaNo ratings yet
- By: Abduljabbar Hamid Jabbar: University of Baghdad-College of Medicine M. B. Ch. BDocument14 pagesBy: Abduljabbar Hamid Jabbar: University of Baghdad-College of Medicine M. B. Ch. BXena XenaNo ratings yet
- FCPS SurgeryDocument106 pagesFCPS Surgeryrehan hayderNo ratings yet
- Obstructive Sleep Apnea Case Study: Shirin Shafazand, MD, MS Neomi Shah, MD August 2014Document15 pagesObstructive Sleep Apnea Case Study: Shirin Shafazand, MD, MS Neomi Shah, MD August 2014Vimal NishadNo ratings yet
- Chapter 15: Health Assessment and Physical ExaminationDocument38 pagesChapter 15: Health Assessment and Physical ExaminationGladz C CadaguitNo ratings yet
- POT Lesson PlanDocument4 pagesPOT Lesson PlanMichelle ThereseNo ratings yet
- Clinical Handbook of Insomnia (2017)Document292 pagesClinical Handbook of Insomnia (2017)jcholguin.conciencia4100100% (8)
- Motor Neurone Disease: J GormallyDocument14 pagesMotor Neurone Disease: J GormallyBambang SutrisnoNo ratings yet
- EMPANELEDHOSPITALLISTDocument12 pagesEMPANELEDHOSPITALLISTVsknNo ratings yet
- Exercise 2Document6 pagesExercise 2Gwyneth Marie DayaganNo ratings yet
- Scale For Ranking Health Conditions and Problems According To PrioritiesDocument9 pagesScale For Ranking Health Conditions and Problems According To Prioritiescnvfiguracion100% (1)
- Safe Working EnvironmentDocument16 pagesSafe Working EnvironmentDawud AsnakewNo ratings yet
- JaundiceDocument4 pagesJaundiceTaleb AbjiNo ratings yet
- Upper GI Drugs (Pod Pharm 2023, Thatcher)Document39 pagesUpper GI Drugs (Pod Pharm 2023, Thatcher)8jm6dhjdcpNo ratings yet
- Community Nursing Care PlanDocument6 pagesCommunity Nursing Care Plantansincos93% (14)
- DrainsDocument19 pagesDrainspaulineNo ratings yet
- Ipcrf TemplateDocument57 pagesIpcrf TemplateJeng SanchezNo ratings yet
- The European Guideline On Management of Major Bleeding and Coagulopathy Following Trauma Sixth Edition (2023) PDFDocument1 pageThe European Guideline On Management of Major Bleeding and Coagulopathy Following Trauma Sixth Edition (2023) PDFMatheus LiraNo ratings yet
- "The Effects of Interpersonal Crime On Victims": Reporter: Michel O. Espinosa Bs-Psychology Code: 3175Document6 pages"The Effects of Interpersonal Crime On Victims": Reporter: Michel O. Espinosa Bs-Psychology Code: 3175Ezri Mariveles Coda Jr.No ratings yet
- Child Guidance ClinicDocument22 pagesChild Guidance ClinicBinal Joshi67% (3)
- Case Study Endometrial PDFDocument17 pagesCase Study Endometrial PDFapi-279886264No ratings yet
- 2012 OiteDocument256 pages2012 Oiteaddison wood100% (4)
- CBD Spinal Infection: by Neoh Zhong NingDocument26 pagesCBD Spinal Infection: by Neoh Zhong NingJoni NeohNo ratings yet
- Day 1 - GynacologyDocument204 pagesDay 1 - GynacologyTingting GeNo ratings yet
- Integumentary System: Burns and Wounds Clinical Case Study: Degree Burn Depth in Dermal Layers Description ImageDocument6 pagesIntegumentary System: Burns and Wounds Clinical Case Study: Degree Burn Depth in Dermal Layers Description ImageGabrielitoNo ratings yet
- Funda DemoDocument10 pagesFunda DemoariadnebabasaNo ratings yet