You might also like
- Faraway WanderersDocument592 pagesFaraway WanderersIsak100% (8)
- Physical Science PDFDocument304 pagesPhysical Science PDFBecca Marie Aled Rolf63% (8)
- Eight Treasures Trousseau PDFDocument1,099 pagesEight Treasures Trousseau PDFSrutokirti Sen100% (1)
- History of Central Market Kuala LumpurDocument7 pagesHistory of Central Market Kuala LumpurXerra ZaraNo ratings yet
- Wang 2019Document10 pagesWang 2019AdeNo ratings yet
- 1 s2.0 S0025326X22007433 MainDocument10 pages1 s2.0 S0025326X22007433 Mainmax tukanaNo ratings yet
- Ingestion of Microplastics by Natural Zooplankton Groups in The Northern SouthDocument8 pagesIngestion of Microplastics by Natural Zooplankton Groups in The Northern SouthMARIBEL DELGADONo ratings yet
- Phytoplankton Composition of Different Fresh Waterbodies of BhopalDocument5 pagesPhytoplankton Composition of Different Fresh Waterbodies of BhopalIJRASETPublicationsNo ratings yet
- Jurnal 2Document8 pagesJurnal 2yut jombang22No ratings yet
- Akhbarizadeh 2020Document11 pagesAkhbarizadeh 2020jhellianeteves14No ratings yet
- Individual and Combined Effects of Amoxicillin and Carbamazepine To The Marine Copepod Tigriopus FulvusDocument10 pagesIndividual and Combined Effects of Amoxicillin and Carbamazepine To The Marine Copepod Tigriopus FulvusFebrina FadhillahNo ratings yet
- Artigo Microplastico 11-10 Docx InglêsDocument22 pagesArtigo Microplastico 11-10 Docx InglêsRayane ReisNo ratings yet
- Environment International: A A B B ADocument8 pagesEnvironment International: A A B B AMeryl QuinteroNo ratings yet
- Warna Mikroplastik Di Mangrove Situbondo & Transparan (Yona)Document9 pagesWarna Mikroplastik Di Mangrove Situbondo & Transparan (Yona)Lina IdamatusNo ratings yet
- 2020 - Buathong - Accumulation of Microplastics in Zooplankton From Chonburi Province, The Upper Gulf of ThailandDocument12 pages2020 - Buathong - Accumulation of Microplastics in Zooplankton From Chonburi Province, The Upper Gulf of ThailandLim Kok PingNo ratings yet
- Bioremediation of Fishpond Effluent and Production of Microalgae For AnDocument12 pagesBioremediation of Fishpond Effluent and Production of Microalgae For AnPriscila Ascencio HenryNo ratings yet
- Environmental PollutionDocument8 pagesEnvironmental PollutionBrigita de BrillarNo ratings yet
- Effects of Microplastic Fibers On Lates Calcarifer Juveniles-AccumulationDocument9 pagesEffects of Microplastic Fibers On Lates Calcarifer Juveniles-Accumulationmegha deyNo ratings yet
- Chen 2018Document40 pagesChen 2018vyvie89No ratings yet
- Akoueson 2020Document10 pagesAkoueson 2020jhellianeteves14No ratings yet
- Microplastic Pollution in Coastal Ecosystem Off Mumbai Coast India Udai Ram Gurjar K A Martin Xavier Satya Prakash Shukla Ashok Kumar Jaiswar Geetanjali Deshmukhe Binaya Bhusan Nayak Full ChapterDocument35 pagesMicroplastic Pollution in Coastal Ecosystem Off Mumbai Coast India Udai Ram Gurjar K A Martin Xavier Satya Prakash Shukla Ashok Kumar Jaiswar Geetanjali Deshmukhe Binaya Bhusan Nayak Full Chapterbrian.collier318100% (17)
- Passability of Chironomid Larvae in Granular ActivDocument7 pagesPassability of Chironomid Larvae in Granular ActivimmufaidatuNo ratings yet
- Heirina 2021 IOP Conf. Ser. Earth Environ. Sci. 744 012026Document13 pagesHeirina 2021 IOP Conf. Ser. Earth Environ. Sci. 744 012026Anna HeirinaNo ratings yet
- Micro PlasticDocument8 pagesMicro PlasticSofia AlviNo ratings yet
- Microplastics Ingestion by Fish in The Pangandaran Bay, IndonesiaDocument9 pagesMicroplastics Ingestion by Fish in The Pangandaran Bay, IndonesiaCianjur RudyNo ratings yet
- Bartonetal 2020BiocontrolofAEFWDocument7 pagesBartonetal 2020BiocontrolofAEFWpawanhv1454No ratings yet
- Ecological Indicators: A B B A ADocument11 pagesEcological Indicators: A B B A ARENZO HERNAN TURPO AROQUIPANo ratings yet
- Journal Pre-Proof: Environmental PollutionDocument37 pagesJournal Pre-Proof: Environmental PollutionPhuping SucharitakulNo ratings yet
- 2024 Arabia PeixeDocument8 pages2024 Arabia PeixeSelma maria de arruda silvaNo ratings yet
- Environmental EngineeringDocument7 pagesEnvironmental EngineeringVeera KanapatyNo ratings yet
- Boyero Et Al. 2020. Microplastics Impair Amphibian Survival, Body Condition and FunctionDocument7 pagesBoyero Et Al. 2020. Microplastics Impair Amphibian Survival, Body Condition and Functionprevencionista servimanefNo ratings yet
- Spatial and Species Variations of Bacterial CommunDocument17 pagesSpatial and Species Variations of Bacterial CommunhiuNo ratings yet
- EFFECT OF MICROPLASTIC ON GREEN MUSSEL Perna Viridis Experimental ApproachDocument6 pagesEFFECT OF MICROPLASTIC ON GREEN MUSSEL Perna Viridis Experimental ApproachKhusnul YaqinNo ratings yet
- 2024 Pakistan Impac HumanoDocument16 pages2024 Pakistan Impac HumanoSelma maria de arruda silvaNo ratings yet
- 1 s2.0 S2352513422002010 MainDocument11 pages1 s2.0 S2352513422002010 MainLuzby María Jiménez CastilloNo ratings yet
- Shayesteh Et Al 2022Document10 pagesShayesteh Et Al 2022Omotola MartinsNo ratings yet
- The Gut Microbiota of Marine Fish: A Historical OverviewDocument17 pagesThe Gut Microbiota of Marine Fish: A Historical OverviewSusila AhmadNo ratings yet
- Aquaculture: A A A A A B B ADocument8 pagesAquaculture: A A A A A B B AJeisson OmarNo ratings yet
- Fmicb 13 907703Document9 pagesFmicb 13 907703Veni DarmaNo ratings yet
- Li 2018Document7 pagesLi 2018Mae MartinezNo ratings yet
- Effects of Microplastics in Marine LifeDocument6 pagesEffects of Microplastics in Marine Lifealexmaximo186No ratings yet
- MésoplanctonsDocument34 pagesMésoplanctonserraiouiNo ratings yet
- Structure and Diversity of Zooplankton Community in Taabo Reservoir (Cote Divoire)Document17 pagesStructure and Diversity of Zooplankton Community in Taabo Reservoir (Cote Divoire)IJAR JOURNALNo ratings yet
- Microplastic Pollution in Water and Fish Samples Around Nanxun Reef inDocument7 pagesMicroplastic Pollution in Water and Fish Samples Around Nanxun Reef inMARIBEL DELGADONo ratings yet
- Ma 2021Document12 pagesMa 2021Essekhyr HassanNo ratings yet
- Effects of Stocking Density On The Growth and Flesh Quality of Rainbow Trout (Oncorhynchus Mykiss) Reared in A Low-Tech Aquaponic SystemDocument40 pagesEffects of Stocking Density On The Growth and Flesh Quality of Rainbow Trout (Oncorhynchus Mykiss) Reared in A Low-Tech Aquaponic Systemjeisson osorioNo ratings yet
- Environmental PollutionDocument13 pagesEnvironmental Pollutionrishabh shuklaNo ratings yet
- Marine Environmental Research: Carlo Giacomo Avio, Stefania Gorbi, Francesco RegoliDocument9 pagesMarine Environmental Research: Carlo Giacomo Avio, Stefania Gorbi, Francesco RegoliEletuzaFariasNo ratings yet
- Drivers of Litter Ingestion by Sea Turtles Three Decades of Empirical DataDocument13 pagesDrivers of Litter Ingestion by Sea Turtles Three Decades of Empirical DataDavid Rodriguez FernandezNo ratings yet
- Artikel 12Document8 pagesArtikel 12Sukma Bening fadliNo ratings yet
- Comparative Study On Phytoplankton Community in Two Newly Dug Ponds in Institut Teknologi SumateraDocument7 pagesComparative Study On Phytoplankton Community in Two Newly Dug Ponds in Institut Teknologi SumateraAhmad JalaludinNo ratings yet
- Kim Et Al. 2017 Bioaccumulation and Elimination E Coli Bacteriophages by Ascidians and BivalvesDocument9 pagesKim Et Al. 2017 Bioaccumulation and Elimination E Coli Bacteriophages by Ascidians and BivalvesRayane SorrentinoNo ratings yet
- Srisunont 2015Document9 pagesSrisunont 2015akosi gantNo ratings yet
- First Bloom BenthicDocument10 pagesFirst Bloom BenthicVictor A. Cervantes-UrietaNo ratings yet
- Jurnal PCA IIDocument12 pagesJurnal PCA IIbiotropic biotropicNo ratings yet
- 2019 - Wang - The Ecotoxicological Effects of Microplastics On Aquatic Food Web, From Primary Producer To Human - A ReviewDocument8 pages2019 - Wang - The Ecotoxicological Effects of Microplastics On Aquatic Food Web, From Primary Producer To Human - A ReviewLim Kok PingNo ratings yet
- Investigating On The Toxicity and Bio Magnification Poten 2023 Marine PollutDocument10 pagesInvestigating On The Toxicity and Bio Magnification Poten 2023 Marine Pollutdika wahyudi purbaNo ratings yet
- Aquaculture 524 (2020) 735277Document11 pagesAquaculture 524 (2020) 735277Jeisson OmarNo ratings yet
- Microplastics Monitoring in Marine Environment IndonesiaDocument6 pagesMicroplastics Monitoring in Marine Environment IndonesianoviNo ratings yet
- Jurnal 1Document26 pagesJurnal 1Rian Dani TumanggorNo ratings yet
- 2021 - Lopez-Martinez - Overview of Global Status of Plastic Presence in Marine VertebratesDocument27 pages2021 - Lopez-Martinez - Overview of Global Status of Plastic Presence in Marine VertebratesLim Kok PingNo ratings yet
- Microplastics in Seawater and ZooplanktonDocument12 pagesMicroplastics in Seawater and ZooplanktonEdwin Salcedo QuiñonesNo ratings yet
- In Vitro Evaluation of Binding of Fish Mucus by Nanoparticles Induce Oxidative Stress On Nile TilapiaDocument14 pagesIn Vitro Evaluation of Binding of Fish Mucus by Nanoparticles Induce Oxidative Stress On Nile TilapiaCentral Asian StudiesNo ratings yet
- Plants From Pitlakes: An inventory of plants from the pitlakes of Eastern Coalfields, IndiaFrom EverandPlants From Pitlakes: An inventory of plants from the pitlakes of Eastern Coalfields, IndiaNo ratings yet
- LM Ucsp Grade11Document314 pagesLM Ucsp Grade11Rhon T. Bergado78% (23)
- EntrepreneurshipDocument191 pagesEntrepreneurshipWacks Venzon50% (2)
- Business Finance TGDocument339 pagesBusiness Finance TGCharisse Dianne P. Andoy100% (2)
- WRB QuizDocument38 pagesWRB QuizMarkKevinAtendidoVidarNo ratings yet
- The Potential of The Coelomic Fluid in Sea CucumbeDocument6 pagesThe Potential of The Coelomic Fluid in Sea CucumbeChristine Joy SulibNo ratings yet
- Different Types of Errors: Blunders Gross Errors Measurement ErrorsDocument1 pageDifferent Types of Errors: Blunders Gross Errors Measurement ErrorsChristine Joy SulibNo ratings yet
- Microplastics ThreatDocument10 pagesMicroplastics ThreatChristine Joy SulibNo ratings yet
- Research AssignmentDocument4 pagesResearch AssignmentChristine Joy SulibNo ratings yet
- Geographical Profiling of Accessibility and MobilityDocument7 pagesGeographical Profiling of Accessibility and MobilityChristine Joy SulibNo ratings yet
- Sea Cucumber Sea SurfaceDocument11 pagesSea Cucumber Sea SurfaceChristine Joy SulibNo ratings yet
- Sea Cucumber Heavy MetalsDocument8 pagesSea Cucumber Heavy MetalsChristine Joy SulibNo ratings yet
- Seacucumber in ChinaDocument30 pagesSeacucumber in ChinaChristine Joy SulibNo ratings yet
- Coastal Rsources - SiquijorDocument9 pagesCoastal Rsources - SiquijorChristine Joy SulibNo ratings yet
- Geographical Profiling of Accessibility and MobilityDocument7 pagesGeographical Profiling of Accessibility and MobilityChristine Joy SulibNo ratings yet
- Js Promenade Cotillion Batch I Line IDocument6 pagesJs Promenade Cotillion Batch I Line IChristine Joy SulibNo ratings yet
- Hypoglycemic 1Document5 pagesHypoglycemic 1Christine Joy SulibNo ratings yet
- Weekly Diary: Sibonga National High SchoolDocument2 pagesWeekly Diary: Sibonga National High SchoolChristine Joy SulibNo ratings yet
- Automated Attendance Monitoring System Using Android PlatformDocument4 pagesAutomated Attendance Monitoring System Using Android PlatformChristine Joy SulibNo ratings yet
- Angiogenic StudyDocument6 pagesAngiogenic StudyChristine Joy SulibNo ratings yet
- Angiogenic 2Document6 pagesAngiogenic 2Christine Joy SulibNo ratings yet
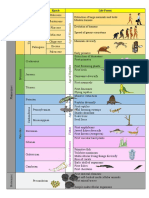
- Eon Era Period Epoch Life FormsDocument2 pagesEon Era Period Epoch Life FormsChristine Joy SulibNo ratings yet
- Coronavirus Disease 2019: An Essay Presented To The Faculty of Sibonga National High SchoolDocument6 pagesCoronavirus Disease 2019: An Essay Presented To The Faculty of Sibonga National High SchoolChristine Joy Sulib100% (1)
- Eon Era Period Epoch Life FormsDocument2 pagesEon Era Period Epoch Life FormsChristine Joy SulibNo ratings yet
- Js Promenade Cotillion Batch I Line IDocument6 pagesJs Promenade Cotillion Batch I Line IChristine Joy SulibNo ratings yet
- Angiogenic StudyDocument6 pagesAngiogenic StudyChristine Joy SulibNo ratings yet
- Table of ContentsDocument2 pagesTable of ContentsChristine Joy SulibNo ratings yet
- Integrated Chinese Level 2 Part 2 Workbook 0Document200 pagesIntegrated Chinese Level 2 Part 2 Workbook 0Kexin UNo ratings yet
- 100 Cities and Their NicknamesDocument4 pages100 Cities and Their NicknamesCharlotte Pacete100% (1)
- Borneo Cross-Cultural Dusun Tatana EthnicDocument3 pagesBorneo Cross-Cultural Dusun Tatana Ethnicplsse 2022No ratings yet
- Qingming Festival (Tomb-Sweeping Day)Document4 pagesQingming Festival (Tomb-Sweeping Day)Momin KhanNo ratings yet
- 听力原文和参考答案 - 副本Document6 pages听力原文和参考答案 - 副本JayNo ratings yet
- PastTime WorksheetDocument16 pagesPastTime Worksheetismail kayaNo ratings yet
- 07 De2Document5 pages07 De2amaterasu.sendaiNo ratings yet
- Kelo Nde AsuDocument3 pagesKelo Nde AsuMr SuyonoNo ratings yet
- Son of The Flying Tiger (K'ing Kung-Fu #1)Document182 pagesSon of The Flying Tiger (K'ing Kung-Fu #1)DWCasonNo ratings yet
- Najua ABCh L2 - Swahili KIDS BookDocument18 pagesNajua ABCh L2 - Swahili KIDS Bookkimanijane11No ratings yet
- 8 Extraordinary Qi VesselsDocument6 pages8 Extraordinary Qi VesselsRiccardo Lautizi100% (1)
- Dissertation Upon Roast Pig AnalysisDocument8 pagesDissertation Upon Roast Pig AnalysisCustomWrittenPapersSingapore100% (1)
- Revision For The First Term: B. Growth B. Think C. EthnicDocument7 pagesRevision For The First Term: B. Growth B. Think C. EthnicHuyền Phạm Thị NgọcNo ratings yet
- Vision A1plusDocument11 pagesVision A1plusIlda ÇibukuNo ratings yet
- Wandering in Northern ChinaDocument628 pagesWandering in Northern ChinaJohn LewisNo ratings yet
- 1001 1500 of 2500 Kanji 500 Entries - CompressDocument17 pages1001 1500 of 2500 Kanji 500 Entries - CompressRawanbadaweiNo ratings yet
- Liste New HSK 1 A 4 Pour MemochinoisDocument65 pagesListe New HSK 1 A 4 Pour MemochinoisPascal DupontNo ratings yet
- Industrial Statistics 7374Document258 pagesIndustrial Statistics 7374Rabin BhandariNo ratings yet
- HSK Study Materials Overview: Total Package Deal Now!Document31 pagesHSK Study Materials Overview: Total Package Deal Now!ManuelNo ratings yet
- Early Fifteenth Century Travels in The East - Nicolò De' Conti of VeniceDocument8 pagesEarly Fifteenth Century Travels in The East - Nicolò De' Conti of VeniceanjanaNo ratings yet
- Young People's Ghost Stories in Hong Kong: JosephboscoDocument23 pagesYoung People's Ghost Stories in Hong Kong: JosephboscoRobert MariasiNo ratings yet
- Celadon: ENG 0103-Test: Unit 4: Local Handicrafts Reading Focus: Scanning TechniqueDocument4 pagesCeladon: ENG 0103-Test: Unit 4: Local Handicrafts Reading Focus: Scanning TechniqueMohamed AlthibiNo ratings yet
- ĐÊ ÔN TẬP HK 1Document6 pagesĐÊ ÔN TẬP HK 1Hồng TrươngNo ratings yet
- Đề Cương Ôn Tập HKI - K8 2020 - 2021Document6 pagesĐề Cương Ôn Tập HKI - K8 2020 - 2021Bích Ngọc ĐặngNo ratings yet
- HSK 3 Vocabulary ListDocument27 pagesHSK 3 Vocabulary ListAnh Thế HoàngNo ratings yet
- Chinese Wedding TraditionsDocument11 pagesChinese Wedding TraditionsCatalinaBrasoveanuNo ratings yet
- New HSK 2 Vocab List PDFDocument18 pagesNew HSK 2 Vocab List PDFAkukuluNo ratings yet