You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5807)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Bashar - 4th Dimension Vrs 4 DensityDocument2 pagesBashar - 4th Dimension Vrs 4 DensityMartin Spyyd Přidal100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Design River Intake TowerDocument24 pagesDesign River Intake TowerDikshant SinghNo ratings yet
- SECOND SUMMATIVE TEST IN SCIENCE 9 q1Document3 pagesSECOND SUMMATIVE TEST IN SCIENCE 9 q1Anna Mae A. Pamonag50% (2)
- Success of Geothermal Wells: A Global StudyDocument80 pagesSuccess of Geothermal Wells: A Global StudyIFC Sustainability100% (1)
- Mathematics in The Modern World Week1 10 QuizzesMidterm PDFDocument11 pagesMathematics in The Modern World Week1 10 QuizzesMidterm PDFJosiah Bebing100% (1)
- CV LabDocument9 pagesCV LabtworedpartyhatsNo ratings yet
- Static Equipment in Oil and Gas IndustryDocument93 pagesStatic Equipment in Oil and Gas IndustryRaghavanNo ratings yet
- Example of Frequency DistributionDocument3 pagesExample of Frequency DistributionJansen Alonzo Bordey80% (5)
- Sample Exam 1 PDFDocument6 pagesSample Exam 1 PDFGirmaye Haile Gebremikael100% (1)
- StockMarket Forecasting Using Hidden Markov Model A New ApproachDocument5 pagesStockMarket Forecasting Using Hidden Markov Model A New Approachandreip21No ratings yet
- The Ecology of Dunaliella in High-Salt Environments: Review Open AccessDocument8 pagesThe Ecology of Dunaliella in High-Salt Environments: Review Open AccessAmmar BouzouerNo ratings yet
- Boukhris-Bouhachem Et Al. 2007 Aphid (Hemiptera Aphidoidea) Diversity in Tunisia in Reionlation With Potato Product PDFDocument9 pagesBoukhris-Bouhachem Et Al. 2007 Aphid (Hemiptera Aphidoidea) Diversity in Tunisia in Reionlation With Potato Product PDFAmmar BouzouerNo ratings yet
- Biomass Recovery and Lipid Extraction Processes For Microalgae Biofuels PDFDocument21 pagesBiomass Recovery and Lipid Extraction Processes For Microalgae Biofuels PDFAmmar BouzouerNo ratings yet
- Bicarbonate-Based Cultivation of Dunaliella Salina For Enhancing Carbon Utilization PDFDocument25 pagesBicarbonate-Based Cultivation of Dunaliella Salina For Enhancing Carbon Utilization PDFAmmar BouzouerNo ratings yet
- Enhancement of Power Quality in Distribution System Using D-Statcom FDocument6 pagesEnhancement of Power Quality in Distribution System Using D-Statcom FNateNo ratings yet
- Chapter 4-6 11-14-Mud Rheology Circuling Systems PumpsDocument117 pagesChapter 4-6 11-14-Mud Rheology Circuling Systems Pumpsbakri OmanNo ratings yet
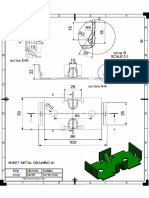
- Siemens NX 10 Sheet Metal TutorialDocument1 pageSiemens NX 10 Sheet Metal TutorialChokri Atef100% (1)
- Business Process BI Services GuideDocument134 pagesBusiness Process BI Services GuideorcatNo ratings yet
- 06D Compressor 2020Document2 pages06D Compressor 2020Syarif HidayatullahNo ratings yet
- ACTOM Outdoor MV Switchgear - SingleDocument8 pagesACTOM Outdoor MV Switchgear - SingleKhashane Willy MohaleNo ratings yet
- Lesson No.1.EXERCISES - MATH FOR ENGINEERS - Function and Algebraic Expression - OCT2021Document4 pagesLesson No.1.EXERCISES - MATH FOR ENGINEERS - Function and Algebraic Expression - OCT2021Audrey MendozaNo ratings yet
- Maximo FormulaDocument9 pagesMaximo FormulaAlbert NguyenNo ratings yet
- Cengage Maths Solutions-Trigonometry Inverse+Trigonometric+FunctionsDocument54 pagesCengage Maths Solutions-Trigonometry Inverse+Trigonometric+Functionskayjayme318No ratings yet
- Final Distribution and Lifespace Products: Price ListDocument66 pagesFinal Distribution and Lifespace Products: Price ListSiddharth SinghNo ratings yet
- Imo 2017Document2 pagesImo 2017sayan paulNo ratings yet
- Institute of Business Administration, Karachi: Schedule - Undergraduate Programs - SPRING SEMESTER 2023 Main CampusDocument6 pagesInstitute of Business Administration, Karachi: Schedule - Undergraduate Programs - SPRING SEMESTER 2023 Main CampusAMIN UDDINNo ratings yet
- LM 78 L 05Document13 pagesLM 78 L 05f7erick-zn5No ratings yet
- Nls Non Linear Summer PDFDocument18 pagesNls Non Linear Summer PDFfreedjackssongwriterNo ratings yet
- Unit One: Introduction To ProgrammingDocument43 pagesUnit One: Introduction To ProgrammingabdiNo ratings yet
- CH 29-Fundamentals of WeldingDocument39 pagesCH 29-Fundamentals of WeldingNasir MaqsoodNo ratings yet
- Dbms 3Document7 pagesDbms 3Panshul SaxenaNo ratings yet
- Software Development Guide 100066 0611 00 enDocument91 pagesSoftware Development Guide 100066 0611 00 enWen HuaNo ratings yet
- Orca Datasheet PDFDocument11 pagesOrca Datasheet PDFaze2010No ratings yet
- Chapter 1 Research Methods-OverviewDocument24 pagesChapter 1 Research Methods-Overviewkassahun meseleNo ratings yet