You might also like
- Dental Research Journal: Microflora and Periodontal DiseaseDocument5 pagesDental Research Journal: Microflora and Periodontal DiseaseNadya PurwantyNo ratings yet
- CALIBR1Document13 pagesCALIBR1Codrin PanainteNo ratings yet
- Periodontal Pathogens and Their Links To Neuroinflammation and NeurodegenerationDocument26 pagesPeriodontal Pathogens and Their Links To Neuroinflammation and NeurodegenerationGeorgi GugicevNo ratings yet
- Prevalence of Six Periodontal Pathogens in Subgingival Samples of Italian Patients With Chronic PeriodontitisDocument8 pagesPrevalence of Six Periodontal Pathogens in Subgingival Samples of Italian Patients With Chronic PeriodontitisDian NoviaNo ratings yet
- Defining The Gut Microbiota in Individuals With Periodontal Diseases: An Exploratory StudyDocument13 pagesDefining The Gut Microbiota in Individuals With Periodontal Diseases: An Exploratory StudyElisabeth TikalakaNo ratings yet
- Pone 0226266Document20 pagesPone 0226266fadhilah RizkiyahNo ratings yet
- Etiologic Role of Root Canal Infection in Apical Periodontitis and Its Relationship With Clinical SymptomatologyDocument29 pagesEtiologic Role of Root Canal Infection in Apical Periodontitis and Its Relationship With Clinical SymptomatologyOmar TalibNo ratings yet
- Microbiology of Periodontal Diseases A ReviewDocument7 pagesMicrobiology of Periodontal Diseases A ReviewghassanNo ratings yet
- Pharmaceutics 14 02740Document18 pagesPharmaceutics 14 02740Robert StryjakNo ratings yet
- Interactions Between Neutrophils and Periodontal Pathogens in Late-Onset PeriodontitisDocument11 pagesInteractions Between Neutrophils and Periodontal Pathogens in Late-Onset PeriodontitisAdriana NoelNo ratings yet
- Antibiotics 10 01298 v3Document15 pagesAntibiotics 10 01298 v3Daniel AtiehNo ratings yet
- Sanz 2004Document14 pagesSanz 2004subbumurugappan94No ratings yet
- Antimicrobial Effectiveness of Cetylpyridinium Chloride and Zinc Chloride-Containing Mouthrinses On Bacteria of Halitosis and Peri-Implant DiseaseDocument7 pagesAntimicrobial Effectiveness of Cetylpyridinium Chloride and Zinc Chloride-Containing Mouthrinses On Bacteria of Halitosis and Peri-Implant DiseasePhuong ThaoNo ratings yet
- 2000 Controlled and Targeted Drug Delivery Strategies Towards Intraperiodontal Pocket DiseasesDocument22 pages2000 Controlled and Targeted Drug Delivery Strategies Towards Intraperiodontal Pocket DiseasesK KNo ratings yet
- Mab200801 01 PDFDocument6 pagesMab200801 01 PDFI Gede MahardikaNo ratings yet
- 1 s2.0 S0882401015001527 MainDocument8 pages1 s2.0 S0882401015001527 MainAndrei AvîrvareiNo ratings yet
- Bacteriology of Most Frequent Oral Anaerobic Infections: S. PiovanoDocument7 pagesBacteriology of Most Frequent Oral Anaerobic Infections: S. PiovanoVinícius LeiteNo ratings yet
- Red Complex Polymicrobial Conglomerate in Oral FloDocument7 pagesRed Complex Polymicrobial Conglomerate in Oral FloDelete AccountNo ratings yet
- Microorganisms 11 00115Document15 pagesMicroorganisms 11 00115Mariana PalaciosNo ratings yet
- Research Article The Periodontal Abscess: Clinical and Microbiological CharacteristicsDocument8 pagesResearch Article The Periodontal Abscess: Clinical and Microbiological CharacteristicsdwiariarthanaNo ratings yet
- 6 Ana PejcicDocument8 pages6 Ana PejcicDermaga_4No ratings yet
- Antibacterial and Anti-Inflammatory Activities of 4-Hydroxycordoin: Potential Therapeutic BenefitsDocument6 pagesAntibacterial and Anti-Inflammatory Activities of 4-Hydroxycordoin: Potential Therapeutic BenefitsVinícius PiantaNo ratings yet
- Lxac 039Document10 pagesLxac 039Cristian PeñaNo ratings yet
- Entamoeba Gingivalis and Trichomonas Tenax In: Periodontal DiseaseDocument12 pagesEntamoeba Gingivalis and Trichomonas Tenax In: Periodontal Diseasefadhilah RizkiyahNo ratings yet
- Aerobic and Anaerobic Bacteria in Subgingival and Supragingival Plaques of Adult Patients With Periodontal DiseaseDocument5 pagesAerobic and Anaerobic Bacteria in Subgingival and Supragingival Plaques of Adult Patients With Periodontal DiseaseRamanda CahyaNo ratings yet
- Wara Aswapati2009 PDFDocument6 pagesWara Aswapati2009 PDFRalph TampubolonNo ratings yet
- (13142143 - Folia Medica) Subgingival Microbiota During Formation of Permanent DentitionDocument7 pages(13142143 - Folia Medica) Subgingival Microbiota During Formation of Permanent DentitionfghdhmdkhNo ratings yet
- Detection of Trichomonas Tenax (T. Tenax) in Bad Oral Hygiene Saudi Patients in Taif CityDocument3 pagesDetection of Trichomonas Tenax (T. Tenax) in Bad Oral Hygiene Saudi Patients in Taif CityInternational Educational Applied Scientific Research Journal (IEASRJ)No ratings yet
- Periodontal Diseases As Bacterial Infection: Bascones Martinez A Figuero Ruiz EDocument8 pagesPeriodontal Diseases As Bacterial Infection: Bascones Martinez A Figuero Ruiz EWidaRostinaNo ratings yet
- ENDO PerioDocument39 pagesENDO PerioapbhalodiNo ratings yet
- Advanced Caries Microbiota in Teeth With Irreversible PulpitisDocument6 pagesAdvanced Caries Microbiota in Teeth With Irreversible PulpitisAbel PelaezNo ratings yet
- 10 1111@jre 12721 PDFDocument7 pages10 1111@jre 12721 PDFSam Bradley DavidsonNo ratings yet
- The Cytokine Network Involved in The Host Immune Response To PeriodontitisDocument13 pagesThe Cytokine Network Involved in The Host Immune Response To PeriodontitisGeorgi GugicevNo ratings yet
- V37n3a02 en IdDocument8 pagesV37n3a02 en IdIip RyanNo ratings yet
- Etiology and Microbiology of Periodontal Diseasea PDFDocument7 pagesEtiology and Microbiology of Periodontal Diseasea PDFlalalalaNo ratings yet
- Current Concepts in The Management of Periodontitis: Concise Clinical ReviewDocument15 pagesCurrent Concepts in The Management of Periodontitis: Concise Clinical ReviewAlexandru Codrin-IonutNo ratings yet
- 55 233 1 PBDocument7 pages55 233 1 PBNadya PurwantyNo ratings yet
- Assessment of Changes in The Oral Microbiome That Occur in Dogs With Periodontal DiseaseDocument12 pagesAssessment of Changes in The Oral Microbiome That Occur in Dogs With Periodontal DiseaseMaria HernandezNo ratings yet
- Int Endodontic J - 2022 - Siqueira - Present Status and Future Directions Microbiology of Endodontic InfectionsDocument19 pagesInt Endodontic J - 2022 - Siqueira - Present Status and Future Directions Microbiology of Endodontic InfectionsHllerd100% (1)
- Trichomonas Tenax and Periodontal Diseases: A Concise ReviewDocument10 pagesTrichomonas Tenax and Periodontal Diseases: A Concise Reviewnursyahrah fahirahNo ratings yet
- P A P C E F: Athogenesis of Pical Eriodontitis and The Auses of Ndodontic AiluresDocument34 pagesP A P C E F: Athogenesis of Pical Eriodontitis and The Auses of Ndodontic AiluresChristine LomuntadNo ratings yet
- Microorganisms 10 02466Document12 pagesMicroorganisms 10 02466Son LeNo ratings yet
- 2 PBDocument7 pages2 PBHenry MandalasNo ratings yet
- 5.siqueira Microbiologia de Infecciones EndodDocument19 pages5.siqueira Microbiologia de Infecciones EndodFernanda CombaNo ratings yet
- Review Article Porphyromonas Gingivalis Periodontal Infection and ItsDocument11 pagesReview Article Porphyromonas Gingivalis Periodontal Infection and ItsMitzi Carolina SierraNo ratings yet
- How To Diagnose and Treat Periodontal-Endodontic Lesions?: Literature Review ArticleDocument7 pagesHow To Diagnose and Treat Periodontal-Endodontic Lesions?: Literature Review ArticleMadrika YantiNo ratings yet
- Abbondanza Tasca - Mineoka 2008Document7 pagesAbbondanza Tasca - Mineoka 2008CORINNE SCALAMBRANo ratings yet
- Rylevprevalence and Distribution Pathogen WorldwideDocument16 pagesRylevprevalence and Distribution Pathogen WorldwideazyytieNo ratings yet
- Basic Sciences: Home MCQ's DiscussionsDocument10 pagesBasic Sciences: Home MCQ's Discussionsdileep9002392No ratings yet
- Oral Biomarkers in The Diagnosis and Progression of Periodontal PDFDocument8 pagesOral Biomarkers in The Diagnosis and Progression of Periodontal PDFAmrí David BarbozaNo ratings yet
- Periodontal Disease: Dental Conference 1IIDocument21 pagesPeriodontal Disease: Dental Conference 1IIClover HidesNo ratings yet
- In Vitro Beneficial Effects of Streptococcus Dentisani As PotentialDocument10 pagesIn Vitro Beneficial Effects of Streptococcus Dentisani As PotentialValentina Villanueva nietoNo ratings yet
- Dental PLAQUE: by Dr. Nitika Jain Post Graduate StudentDocument70 pagesDental PLAQUE: by Dr. Nitika Jain Post Graduate StudentJee ArceoNo ratings yet
- Endo-Perio Dilemma A Brief ReviewDocument16 pagesEndo-Perio Dilemma A Brief ReviewZahra Khairiza AnriNo ratings yet
- 2022 - Horvat Aleksijevic - Porphyromonas Gingivalis - Pathogens-11-01173Document19 pages2022 - Horvat Aleksijevic - Porphyromonas Gingivalis - Pathogens-11-01173Ivana V SkrlecNo ratings yet
- Overview of Periodontal Disease, Etiology, Pathogenesis, Diagnosis and Treatment TherapyDocument10 pagesOverview of Periodontal Disease, Etiology, Pathogenesis, Diagnosis and Treatment TherapyNarito NianNo ratings yet
- Management of Aggressive Periodontitis Using Ozonized WaterDocument17 pagesManagement of Aggressive Periodontitis Using Ozonized Wateraramirezp09No ratings yet
- Endo-Perio Dilemma A Brief ReviewDocument10 pagesEndo-Perio Dilemma A Brief ReviewLuisNo ratings yet
- Cats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisFrom EverandCats and Toxoplasma: A Comprehensive Guide to Feline ToxoplasmosisNo ratings yet
- PhenotypeDocument14 pagesPhenotypeDragos CiongaruNo ratings yet
- Integrating Conventional and CAD/CAM Digital Techniques For Establishing Canine Protected Articulation: A Clinical ReportDocument5 pagesIntegrating Conventional and CAD/CAM Digital Techniques For Establishing Canine Protected Articulation: A Clinical ReportDragos CiongaruNo ratings yet
- Yin 2017Document14 pagesYin 2017Odalis CuaspudNo ratings yet
- 14 - Preparations For Periodontally Weakened TeethDocument24 pages14 - Preparations For Periodontally Weakened TeethDragos CiongaruNo ratings yet
- Dsdapp Use For Multidisciplinary Esthetic PlanningDocument9 pagesDsdapp Use For Multidisciplinary Esthetic PlanningDragos CiongaruNo ratings yet
- A Method of Locating The Abutment Screw Access Channel With Cone-Beam Computed Tomography and A 3d-Printed Drilling GuideDocument4 pagesA Method of Locating The Abutment Screw Access Channel With Cone-Beam Computed Tomography and A 3d-Printed Drilling GuideDragos CiongaruNo ratings yet
- Articol PivotDocument7 pagesArticol PivotDragos CiongaruNo ratings yet
- Yin 2017Document14 pagesYin 2017Odalis CuaspudNo ratings yet
- Surgical Extrusion: A Dental TechniqueDocument6 pagesSurgical Extrusion: A Dental TechniqueDragos CiongaruNo ratings yet
- Implant Therapy For A Patient With Down Syndrome and Oral Habits: A Clinical ReportDocument5 pagesImplant Therapy For A Patient With Down Syndrome and Oral Habits: A Clinical ReportDragos CiongaruNo ratings yet
- Castelo-Baz2019 Surgical ExtrusionDocument4 pagesCastelo-Baz2019 Surgical Extrusionnithya_sendhilNo ratings yet
- Yin 2017Document14 pagesYin 2017Odalis CuaspudNo ratings yet
- Dsdapp Use For Multidisciplinary Esthetic PlanningDocument9 pagesDsdapp Use For Multidisciplinary Esthetic PlanningDragos CiongaruNo ratings yet
- Use of Intraoral Scanning and 3-Dimensional Printing in The Fabrication of A Removable Partial Denture For A Patient With Limited Mouth OpeningDocument4 pagesUse of Intraoral Scanning and 3-Dimensional Printing in The Fabrication of A Removable Partial Denture For A Patient With Limited Mouth OpeningDragos CiongaruNo ratings yet
- Al-Harbi - 2018 - A Guide To Minimally Invasive Crown Lengthening and Tooth Preparation For Rehabilitating Pink and White AeDocument5 pagesAl-Harbi - 2018 - A Guide To Minimally Invasive Crown Lengthening and Tooth Preparation For Rehabilitating Pink and White AefishribbonNo ratings yet
- Use of Intraoral Scanning and 3-Dimensional Printing in The Fabrication of A Removable Partial Denture For A Patient With Limited Mouth OpeningDocument4 pagesUse of Intraoral Scanning and 3-Dimensional Printing in The Fabrication of A Removable Partial Denture For A Patient With Limited Mouth OpeningDragos CiongaruNo ratings yet
- 2018 OCNAENTJPD Volume 120 Issue 1 July PDFDocument4 pages2018 OCNAENTJPD Volume 120 Issue 1 July PDFhinanaimNo ratings yet
- Integrating Conventional and CAD/CAM Digital Techniques For Establishing Canine Protected Articulation: A Clinical ReportDocument5 pagesIntegrating Conventional and CAD/CAM Digital Techniques For Establishing Canine Protected Articulation: A Clinical ReportDragos CiongaruNo ratings yet
- Sahrmann 2010Document12 pagesSahrmann 2010Dragos CiongaruNo ratings yet
- An Alternative Solution For A Complex Prosthodontic Problem: A Modified Andrews Fixed Dental ProsthesisDocument5 pagesAn Alternative Solution For A Complex Prosthodontic Problem: A Modified Andrews Fixed Dental ProsthesisDragos CiongaruNo ratings yet
- A Method of Locating The Abutment Screw Access Channel With Cone-Beam Computed Tomography and A 3d-Printed Drilling GuideDocument4 pagesA Method of Locating The Abutment Screw Access Channel With Cone-Beam Computed Tomography and A 3d-Printed Drilling GuideDragos CiongaruNo ratings yet
- Castelo-Baz2019 Surgical ExtrusionDocument4 pagesCastelo-Baz2019 Surgical Extrusionnithya_sendhilNo ratings yet
- Prosthetic Management of An Existing Transmandibular Implant: A Clinical ReportDocument5 pagesProsthetic Management of An Existing Transmandibular Implant: A Clinical ReportDragos CiongaruNo ratings yet
- Digital Bar Prototype Technique For Full-Arch Rehabilitation On ImplantsDocument7 pagesDigital Bar Prototype Technique For Full-Arch Rehabilitation On ImplantsDragos CiongaruNo ratings yet
- Multistage Implantology-Orthodontic-Prosthetic Treatment: A Case ReportDocument10 pagesMultistage Implantology-Orthodontic-Prosthetic Treatment: A Case ReportDragos CiongaruNo ratings yet
- Yin 2017Document14 pagesYin 2017Odalis CuaspudNo ratings yet
- Cortellini Et Al-2018-Journal of Clinical PeriodontologyDocument9 pagesCortellini Et Al-2018-Journal of Clinical PeriodontologyErnestoNo ratings yet
- Prosthetic Management of An Existing Transmandibular Implant: A Clinical ReportDocument5 pagesProsthetic Management of An Existing Transmandibular Implant: A Clinical ReportDragos CiongaruNo ratings yet
- (23915412 - Open Life Sciences) The Effects of Various Mouthwashes On Osteoblast Precursor CellsDocument8 pages(23915412 - Open Life Sciences) The Effects of Various Mouthwashes On Osteoblast Precursor CellsDragos CiongaruNo ratings yet
- Mining Legislation and Mineral Development in ZambiaDocument35 pagesMining Legislation and Mineral Development in ZambiaNathan MwewaNo ratings yet
- Pengisian Tanggal3Document32 pagesPengisian Tanggal3gusrina simamoraNo ratings yet
- Fakultas Kedokteran: Kementerian Pendidikan Dan Kebudayaan Universitas Jenderal SoedirmanDocument1 pageFakultas Kedokteran: Kementerian Pendidikan Dan Kebudayaan Universitas Jenderal SoedirmanChausarPutraBenagilNo ratings yet
- Method Statement For Installation of Telephone ManholeDocument4 pagesMethod Statement For Installation of Telephone ManholemujtiobamaliblNo ratings yet
- Improve Your Medication Management Processes: Advisory ServicesDocument2 pagesImprove Your Medication Management Processes: Advisory ServicesSimarpreet KaurNo ratings yet
- Clinical Cardiology - April 1994 - Wareing - Cardiac Surgery 2nd Edition by John W Kirklin and Brian G Barratt BoyesDocument1 pageClinical Cardiology - April 1994 - Wareing - Cardiac Surgery 2nd Edition by John W Kirklin and Brian G Barratt BoyesOlga KochubeiNo ratings yet
- ResearchDocument70 pagesResearchMagud, Jemverick Jan Leonard O.No ratings yet
- Sweet Dreams KitDocument1 pageSweet Dreams KitPoplacenel Serban DanNo ratings yet
- Canine and Feline Tail Amputation: Clinical TechniquesDocument2 pagesCanine and Feline Tail Amputation: Clinical TechniquesHalwa MulyadiNo ratings yet
- Mhcet Mock Test-2Document29 pagesMhcet Mock Test-2Poonam UpadhyayNo ratings yet
- 1K Solid BasecoatDocument6 pages1K Solid BasecoatjanakaNo ratings yet
- Arrangement of Artificial Teeth: Dr. Enas Abdul-SattarDocument50 pagesArrangement of Artificial Teeth: Dr. Enas Abdul-SattarShashank AadarshNo ratings yet
- Oi 247-2 (En) Rev12 2016 04 21 PDFDocument222 pagesOi 247-2 (En) Rev12 2016 04 21 PDFEhsanNo ratings yet
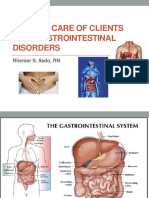
- Nursing of Clients With Gastrointestinal DisordersDocument228 pagesNursing of Clients With Gastrointestinal DisordersLane Mae Magpatoc Noerrot100% (1)
- Technical Requirements in Vitro Diagnostics (IVD)Document4 pagesTechnical Requirements in Vitro Diagnostics (IVD)Raydoon Sadeq100% (1)
- HDFC ERGO General Insurance Company Limited: Claim Form - Part ADocument7 pagesHDFC ERGO General Insurance Company Limited: Claim Form - Part AchakshuNo ratings yet
- Fundamentals of Nursing Transes 3Document4 pagesFundamentals of Nursing Transes 3Louise TorresNo ratings yet
- 5 INGREDIENTS - EbookDocument14 pages5 INGREDIENTS - EbookHeuler KretliNo ratings yet
- Sop of Disinfection, CoatingDocument27 pagesSop of Disinfection, CoatingAhmed Quadri0% (1)
- Andreas Tigor 22010112130144 Lap Kti Bab7Document19 pagesAndreas Tigor 22010112130144 Lap Kti Bab7Zia NajlaNo ratings yet
- Tork Catalogue UkiDocument102 pagesTork Catalogue UkiBirdsNo ratings yet
- Sach Sentence Completion Test InterpretationDocument4 pagesSach Sentence Completion Test Interpretationrupal arora67% (3)
- Biotechnology Assignment SolutionDocument8 pagesBiotechnology Assignment SolutionPiyush RajNo ratings yet
- Queer Identity and Gender-Related Rights in Post-Colonial PhilippinesDocument12 pagesQueer Identity and Gender-Related Rights in Post-Colonial PhilippinesSkord 176No ratings yet
- Al-Okshi Et Al 2019Document9 pagesAl-Okshi Et Al 2019Ayman AlOkshiNo ratings yet
- Ruby NCPDocument3 pagesRuby NCPRuby Jane LaquihonNo ratings yet
- Kepelbagaian Budaya (Individual Assignment)Document6 pagesKepelbagaian Budaya (Individual Assignment)Yusran RosdiNo ratings yet
- Jacobi Aquasorb F23 SDSDocument10 pagesJacobi Aquasorb F23 SDSkhiemNo ratings yet
- The Leadership Effectiveness and Adaptability Descriptor Tool AnalysisDocument4 pagesThe Leadership Effectiveness and Adaptability Descriptor Tool Analysisashlesha_ganatra100% (8)
- Method of Statement For GroutingDocument7 pagesMethod of Statement For GroutingCezar DasiNo ratings yet