You might also like
- Delphinidin Activates NFAT and Induces Cytokine Production via SOCEDocument13 pagesDelphinidin Activates NFAT and Induces Cytokine Production via SOCECristobal ParraNo ratings yet
- Effect of Phosphocreatine and EthylmethyDocument2 pagesEffect of Phosphocreatine and Ethylmethymercuryboss90No ratings yet
- Magnesium Ion Influx Reduces Neuroinflammation in Ab Precursor Protein/presenilin 1 Transgenic Mice by Suppressing The Expression of Interleukin-1bDocument14 pagesMagnesium Ion Influx Reduces Neuroinflammation in Ab Precursor Protein/presenilin 1 Transgenic Mice by Suppressing The Expression of Interleukin-1bkleberttoscanoNo ratings yet
- Jvms 83 997Document7 pagesJvms 83 997Blanca A SerranoNo ratings yet
- An Effect of Protein Kinase C Activation: Induction of Endothelin-1 Expression by GlucoseDocument10 pagesAn Effect of Protein Kinase C Activation: Induction of Endothelin-1 Expression by GlucosePutra Rahmadea UtamiNo ratings yet
- PUBLICADO JCBiochem - 2012Document12 pagesPUBLICADO JCBiochem - 2012Claudia BuitragoNo ratings yet
- Collecting Duct-Specific Knockout of Endothelin-1 Causes Hypertension and Sodium RetentionDocument8 pagesCollecting Duct-Specific Knockout of Endothelin-1 Causes Hypertension and Sodium RetentionMark KoshlandNo ratings yet
- Quimeras en CabrasDocument9 pagesQuimeras en Cabraslauradaza_89No ratings yet
- In Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinDocument6 pagesIn Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinElijah James LegaspiNo ratings yet
- 2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Document9 pages2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Pablo Tapia BatallaNo ratings yet
- Pulmón e Isquemia IntestinalDocument13 pagesPulmón e Isquemia IntestinalOperacion ConocimientoNo ratings yet
- IL27 CardioprotectoraDocument13 pagesIL27 CardioprotectoraAdrián Asael Rodriguez CortesNo ratings yet
- Structure Kestose and NystoseDocument6 pagesStructure Kestose and NystoseJavier MendozaNo ratings yet
- Reversal of Streptozotocin-Induced Hyperglycemia by Transplantation of Pseudoislets Consisting of From Ductal CellsDocument6 pagesReversal of Streptozotocin-Induced Hyperglycemia by Transplantation of Pseudoislets Consisting of From Ductal CellskalwusaidiNo ratings yet
- Trypanosoma congolense Gene ExpressionDocument0 pagesTrypanosoma congolense Gene ExpressionMaísa FonsecaNo ratings yet
- Url-2003 Parvovirus en Neuronas de GatoDocument5 pagesUrl-2003 Parvovirus en Neuronas de GatocristianNo ratings yet
- University of GroningenDocument6 pagesUniversity of GroningenBreixo HarguindeyNo ratings yet
- Macrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin ReceptorsDocument8 pagesMacrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin Receptorsvanessa_murillo_5No ratings yet
- Beta-Cell-Mimetic Designer Cells Provide Closed-Loop Glycemic ControlDocument7 pagesBeta-Cell-Mimetic Designer Cells Provide Closed-Loop Glycemic ControlNando93No ratings yet
- Refolding and Characterization of RecombDocument6 pagesRefolding and Characterization of RecombMobarakNo ratings yet
- Astrocytic Expression of CTMP Following An Excitotoxic Lesion in The Mouse HippocampusDocument8 pagesAstrocytic Expression of CTMP Following An Excitotoxic Lesion in The Mouse HippocampusSergeat18BNo ratings yet
- Liu 2005Document7 pagesLiu 2005Javier Cerda InfanteNo ratings yet
- Ajpendo 90369 2008Document9 pagesAjpendo 90369 2008Maria Virgina Perdomo De Los SantosNo ratings yet
- IFN e VEGF-CDocument11 pagesIFN e VEGF-CThádia AraújoNo ratings yet
- Crocetin Protects Against Cardiac Hypertrophy by Blocking MEK-ERK1/2 Signalling PathwayDocument17 pagesCrocetin Protects Against Cardiac Hypertrophy by Blocking MEK-ERK1/2 Signalling PathwayHugo ARNo ratings yet
- AsdDocument15 pagesAsdRaditya KarunaNo ratings yet
- Cai Et Al. - 2012 - The Heme Oxygenase 1 Inducer (CoPP) Protects Human Cardiac Stem Cells Against Apoptosis Through Activation of The ExDocument17 pagesCai Et Al. - 2012 - The Heme Oxygenase 1 Inducer (CoPP) Protects Human Cardiac Stem Cells Against Apoptosis Through Activation of The Exscribd mendeleyNo ratings yet
- Eneregetski MetabolizamDocument6 pagesEneregetski MetabolizamMarko AdamovicNo ratings yet
- Hepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6Document12 pagesHepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6relbelkasyNo ratings yet
- Gentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsDocument0 pagesGentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsCLPHtheoryNo ratings yet
- Increased Threshold For TCR-Mediated Signaling Controls Self Reactivity of Intraepithelial LymphocytesDocument7 pagesIncreased Threshold For TCR-Mediated Signaling Controls Self Reactivity of Intraepithelial LymphocytesBiosynthesisNo ratings yet
- Primary Neurons Can Enter M-PhaseDocument15 pagesPrimary Neurons Can Enter M-PhaseManuel PachicanoNo ratings yet
- Cross 1987 Catecholamines LympohcytesDocument7 pagesCross 1987 Catecholamines LympohcytesMartin BadovNo ratings yet
- Halloran 2004Document15 pagesHalloran 2004Agustina ParnisariNo ratings yet
- GIVORD 2018 - Activation of the endoplasmic reticulum stress sensor IRE1α by the vaccine adjuvant AS03 contributes to its immunostimulatory propertiesDocument12 pagesGIVORD 2018 - Activation of the endoplasmic reticulum stress sensor IRE1α by the vaccine adjuvant AS03 contributes to its immunostimulatory propertiesJoão Pedro NunesNo ratings yet
- Cdc42 Regulation by Intersectin 2 During Xenopus DevelopmentDocument6 pagesCdc42 Regulation by Intersectin 2 During Xenopus DevelopmentSergeat18BNo ratings yet
- Mechanism of Macrophage-Derived Chemokine Production by KeratinocytesDocument5 pagesMechanism of Macrophage-Derived Chemokine Production by Keratinocytesoscarbio2009No ratings yet
- κB Kinase-dependent κB Activation: M olecules and CellsDocument9 pagesκB Kinase-dependent κB Activation: M olecules and CellsLucas TobingNo ratings yet
- The Ascorbic Acid Transporter SVCT2 Is Expressed in Slow-Twitch Skeletal Muscle WbresDocument10 pagesThe Ascorbic Acid Transporter SVCT2 Is Expressed in Slow-Twitch Skeletal Muscle WbresJorge Eneas DosoliNo ratings yet
- Tian 2016Document13 pagesTian 2016Epiphany SonderNo ratings yet
- Curcumin - ECMDocument7 pagesCurcumin - ECMBABANG GAGAHNo ratings yet
- Phytomedicine: Original ArticleDocument9 pagesPhytomedicine: Original ArticleAnjali MohanNo ratings yet
- Celecoxib y ApoptosisDocument7 pagesCelecoxib y ApoptosisOperacion ConocimientoNo ratings yet
- (Nishikawa Et Al. 2000) Normalizing Mitochondrial Superoxide Production Blocks Three Pathways of Hyperglycaemic DamageDocument4 pages(Nishikawa Et Al. 2000) Normalizing Mitochondrial Superoxide Production Blocks Three Pathways of Hyperglycaemic DamageSie ningsihNo ratings yet
- Jlms 10 310Document7 pagesJlms 10 310Victor RoticivNo ratings yet
- 2010 ER Stress Induction in T CellDocument6 pages2010 ER Stress Induction in T CellRaul ReyesNo ratings yet
- Congress PresentationDocument15 pagesCongress PresentationRahmat AbdullahNo ratings yet
- 1 s2.0 S0014482702955813 MainDocument12 pages1 s2.0 S0014482702955813 MainKevin MaiseyNo ratings yet
- Differential effects of PGE1 and PGE2 on murine myeloid leukemia cellsDocument7 pagesDifferential effects of PGE1 and PGE2 on murine myeloid leukemia cellswindyndaNo ratings yet
- Inducible Immune Regulation Following Autoimmune Disease in The Immune-Privileged EyeDocument7 pagesInducible Immune Regulation Following Autoimmune Disease in The Immune-Privileged EyeDarren J LeeNo ratings yet
- PKD2 eIF2a HMG 2008Document9 pagesPKD2 eIF2a HMG 2008Xing-Zhen ChenNo ratings yet
- Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsDocument7 pagesOverexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsCamilo CabreraNo ratings yet
- 10.Luteolin Suppresses Cisplatin-Induced Apoptosis in Auditory Cells Possible Mediation Through Induction of Heme Oxygenase-1 Expression的副本Document7 pages10.Luteolin Suppresses Cisplatin-Induced Apoptosis in Auditory Cells Possible Mediation Through Induction of Heme Oxygenase-1 Expression的副本Shawn GaoNo ratings yet
- Hypertonic Stress Regulates T Cell Function Via Pannexin-1 Hemichannels and P2X ReceptorsDocument9 pagesHypertonic Stress Regulates T Cell Function Via Pannexin-1 Hemichannels and P2X ReceptorsjavillusNo ratings yet
- Enhancement Effects of Nicotine On Neuro PDFDocument7 pagesEnhancement Effects of Nicotine On Neuro PDFEvelyn Conan HolmesNo ratings yet
- Wang2007 Icaritin NeuroprotectionDocument12 pagesWang2007 Icaritin Neuroprotectioniulia andreeaNo ratings yet
- ArticuloDocument8 pagesArticuloRubi Diaz RomeroNo ratings yet
- 1241 Full PDFDocument13 pages1241 Full PDFMuhammad FaheemNo ratings yet
- Quercetin PankreasDocument7 pagesQuercetin PankreasAlmas TNo ratings yet
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- PMA Up-Regulates The Transcription of Axl by AP-1 Transcription Factor Binding To TRE Sequences Via The MAPK Cascade in Leukaemia CellsDocument19 pagesPMA Up-Regulates The Transcription of Axl by AP-1 Transcription Factor Binding To TRE Sequences Via The MAPK Cascade in Leukaemia CellsWilliam RamirezNo ratings yet
- J. Biol. Chem.-1997-Bennett-8137-40Document5 pagesJ. Biol. Chem.-1997-Bennett-8137-40William RamirezNo ratings yet
- Characterization of Expressed Genes in Kidney Cells of Japanese Flounder Paralichthys Olivaceus Following Treatment With Cona/Pma and LpsDocument8 pagesCharacterization of Expressed Genes in Kidney Cells of Japanese Flounder Paralichthys Olivaceus Following Treatment With Cona/Pma and LpsWilliam RamirezNo ratings yet
- Increased Resistin May Suppress Reactive Oxygen SPDocument11 pagesIncreased Resistin May Suppress Reactive Oxygen SPWilliam RamirezNo ratings yet
- Effect of Antibodies Double Stranded DNA, Purified Samples of With Lupus Erythematosus, The Glomerular Mesangial CellsDocument6 pagesEffect of Antibodies Double Stranded DNA, Purified Samples of With Lupus Erythematosus, The Glomerular Mesangial CellsWilliam RamirezNo ratings yet
- 000337953200010Document10 pages000337953200010William RamirezNo ratings yet
- Passive Fire System Checklist UbblDocument6 pagesPassive Fire System Checklist UbblXinbby BeexinNo ratings yet
- Warm Vs Cold ShockDocument8 pagesWarm Vs Cold ShockmarleeramirezNo ratings yet
- Duarte Et Al - Resin CementsDocument28 pagesDuarte Et Al - Resin Cementsjfrascarolo100% (4)
- Predicting Natural Hazards PowerPointDocument29 pagesPredicting Natural Hazards PowerPointZarlene SierraNo ratings yet
- ETR - Part C - Course SyllabusDocument26 pagesETR - Part C - Course SyllabusDon JubacNo ratings yet
- Technical Data Sheet PDFDocument2 pagesTechnical Data Sheet PDFLuis VilelaNo ratings yet
- Subway 2009Document12 pagesSubway 2009sparklers_mNo ratings yet
- Grade 5. Unit 15Document6 pagesGrade 5. Unit 15Đình ThuậnNo ratings yet
- 14 People V Manalo (Verba Legis)Document15 pages14 People V Manalo (Verba Legis)zeynNo ratings yet
- DBXLH 8585B VTMDocument4 pagesDBXLH 8585B VTMmau_mmx5738No ratings yet
- Animal Names in English and Telugu - 20170912 - 182216545Document17 pagesAnimal Names in English and Telugu - 20170912 - 182216545karimulla goraNo ratings yet
- Cochlear ImplantsDocument53 pagesCochlear ImplantsSovitJungBaralNo ratings yet
- Wrap Book A4Document27 pagesWrap Book A4doscribe100% (1)
- Fertilization to Implantation StagesDocument18 pagesFertilization to Implantation StagesNurulAqilahZulkifliNo ratings yet
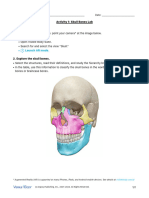
- Lab Manual - Skull Bones - English - Student - Fill inDocument6 pagesLab Manual - Skull Bones - English - Student - Fill inthejaskumar2026No ratings yet
- Recommended Abma & Asme Boiler Water Limits Drum Operating Pressure (Psig) SteamDocument9 pagesRecommended Abma & Asme Boiler Water Limits Drum Operating Pressure (Psig) Steammaoc4vnNo ratings yet
- Online Medicine Industry Competitor AnalysisDocument14 pagesOnline Medicine Industry Competitor Analysispawangadiya1210No ratings yet
- A Study On Satisfaction Level of Employees With Special Reference Textile IndustryDocument12 pagesA Study On Satisfaction Level of Employees With Special Reference Textile Industrysai kiran bade100% (1)
- Mind Power SecretsDocument38 pagesMind Power SecretsIan Faian92% (52)
- Laporan FaalDocument25 pagesLaporan FaalAgnes NathaniaNo ratings yet
- An Integrated Review of Evidence-Based Healthcare Design For Healing Environments Focusing On Longterm Care FacilitiesDocument16 pagesAn Integrated Review of Evidence-Based Healthcare Design For Healing Environments Focusing On Longterm Care FacilitiesAmira EsamNo ratings yet
- Product Brochure-Electronically Controlled Air Dryer-ECA PDFDocument4 pagesProduct Brochure-Electronically Controlled Air Dryer-ECA PDFAnonymous O0T8aZZNo ratings yet
- Argumentative EssayDocument5 pagesArgumentative Essayapi-538443988No ratings yet
- ICICI Pru IProtect Smart Illustrated BrochureDocument56 pagesICICI Pru IProtect Smart Illustrated Brochuresoubhadra nagNo ratings yet
- Understanding Immunomodulatory DrugsDocument4 pagesUnderstanding Immunomodulatory DrugsMark Russel Sean LealNo ratings yet
- BInggrisDocument19 pagesBInggrisanwar318No ratings yet
- Structural Notes and Specifications for ConcreteDocument1 pageStructural Notes and Specifications for Concreteroger100% (2)
- Manual Book IOG ConventionDocument17 pagesManual Book IOG ConventionTaufiq MaulanaNo ratings yet
- Chapter 13 - Chain and SprocketsDocument78 pagesChapter 13 - Chain and SprocketsSugeng Ciko DarmintoNo ratings yet
- Topic - 2015 Quality MethotdDocument12 pagesTopic - 2015 Quality MethotdclaudiaNo ratings yet
- Passenger Ship SubdivisionDocument6 pagesPassenger Ship SubdivisionPunit Pandey100% (2)