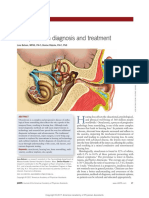
You might also like
- Noise and the Brain: Experience Dependent Developmental and Adult PlasticityFrom EverandNoise and the Brain: Experience Dependent Developmental and Adult PlasticityNo ratings yet
- Noise Induced Loss of Hair Cells and Cochlear Synaptopathy Are Mediated by The Activation of AMPKDocument14 pagesNoise Induced Loss of Hair Cells and Cochlear Synaptopathy Are Mediated by The Activation of AMPKSaifuddin HaswareNo ratings yet
- Acoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inDocument11 pagesAcoustic and Optoacoustic Stimulations in Auditory Brainstem Response Test inabsabravo4No ratings yet
- Otorhinolaryngology: World Journal ofDocument14 pagesOtorhinolaryngology: World Journal ofLeonardoMartínezNo ratings yet
- The Role of Antioxidants in Protection From Impulse NoiseDocument13 pagesThe Role of Antioxidants in Protection From Impulse NoiseStefan CanicNo ratings yet
- Assessment of Hidden Hearing Loss in Noise Exposed WorkersDocument14 pagesAssessment of Hidden Hearing Loss in Noise Exposed WorkersIJAR JOURNALNo ratings yet
- Presbycusis 181116131215 PDFDocument66 pagesPresbycusis 181116131215 PDFWael ShamyNo ratings yet
- Trauma Acoustic JournalDocument6 pagesTrauma Acoustic JournalLannydchandraNo ratings yet
- Ocupational Hearing LossDocument38 pagesOcupational Hearing LossNIKOLAOS CHATZAKISNo ratings yet
- International Journal of Pediatric OtorhinolaryngologyDocument7 pagesInternational Journal of Pediatric OtorhinolaryngologyIlham TaufanNo ratings yet
- Cochlear Dysfunction With Acoustic Trauma in Fire Shooting TrainingDocument5 pagesCochlear Dysfunction With Acoustic Trauma in Fire Shooting TrainingDesywinNo ratings yet
- Ijms 24 02947 v3Document15 pagesIjms 24 02947 v3FuninganaNo ratings yet
- Schwannoma Vestibular TinnitusDocument6 pagesSchwannoma Vestibular TinnitusThomasMáximoMancinelliRinaldoNo ratings yet
- 585 600 HellemanDocument17 pages585 600 HellemanBudi Darmawan DiswanNo ratings yet
- Er Noise Exposure Whitepaper 2014 PDFDocument8 pagesEr Noise Exposure Whitepaper 2014 PDFrpercy01No ratings yet
- Auditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case ReportsDocument29 pagesAuditory Neuropathy Spectrum Disorders: From Diagnosis To Treatment: Literature Review and Case Reportschipta lestariNo ratings yet
- Prog Oftalmo 1 2019Document6 pagesProg Oftalmo 1 2019FerviferrazNo ratings yet
- Effects of Noise in Animals. Turner2005Document28 pagesEffects of Noise in Animals. Turner2005Muzi Valentina RojasNo ratings yet
- Research Article: C M. T, W L, D M F, R MDocument8 pagesResearch Article: C M. T, W L, D M F, R MArizha IrmaNo ratings yet
- The Physiology of Cochlear Presbycusis: Richard A. SchmiedtDocument31 pagesThe Physiology of Cochlear Presbycusis: Richard A. Schmiedtemilia candraNo ratings yet
- Longenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMDocument12 pagesLongenecker-Galazyuk2011 Article DevelopmentOfTinnitusInCBACaJMannaNo ratings yet
- CURRENT Diagnosis & Treatment in Otolaryngology-Head & Neck Surgery, 3eDocument3 pagesCURRENT Diagnosis & Treatment in Otolaryngology-Head & Neck Surgery, 3eStephanie Vania EmbangNo ratings yet
- Rehab - Hi (New)Document3 pagesRehab - Hi (New)devNo ratings yet
- Danesh 2018Document16 pagesDanesh 2018gemapesfNo ratings yet
- International Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanDocument11 pagesInternational Journal of Developmental Neuroscience: S.L. Uran, M.E. Gómez-Casati, L.R. GuelmanWoody WoodyNo ratings yet
- Auditory and Non-Auditory Eff Ects of Noise On HealthDocument8 pagesAuditory and Non-Auditory Eff Ects of Noise On HealthLeonardoMartínezNo ratings yet
- Sensorineural Hearing Loss Associated With Occupational Noise Exposure: Effects of Age-CorrectionsDocument11 pagesSensorineural Hearing Loss Associated With Occupational Noise Exposure: Effects of Age-CorrectionsLembah BarokahNo ratings yet
- 405 2013 Article 2849Document8 pages405 2013 Article 2849nisawidiyawardaniNo ratings yet
- OAE Pediatrician BookletDocument22 pagesOAE Pediatrician BookletKalpana ParajuliNo ratings yet
- Fuente 2012Document8 pagesFuente 2012Leydi PilarNo ratings yet
- How To Reverse Hearing Loss and TinnitusDocument24 pagesHow To Reverse Hearing Loss and TinnitusMark Sloan100% (4)
- Restoration of Hearing LossDocument10 pagesRestoration of Hearing LossZhungli QuanNo ratings yet
- Noise Induces Oxidative Stress in RatDocument5 pagesNoise Induces Oxidative Stress in RatflorensiaNo ratings yet
- Frequency Responses of Headphones Modulate Alpha Brain OscillationsDocument10 pagesFrequency Responses of Headphones Modulate Alpha Brain Oscillationsnataly Delgado MontecinosNo ratings yet
- Brozoski2012 Article BilateralDorsalCochlearNucleusDocument12 pagesBrozoski2012 Article BilateralDorsalCochlearNucleusLanny Dwi ChandraNo ratings yet
- Occupational Noise Exposure and Risk For Noise-Induced Hearing Loss Due To Temporal Bone DrillingDocument7 pagesOccupational Noise Exposure and Risk For Noise-Induced Hearing Loss Due To Temporal Bone DrillingAnanda Chaerunnisa PattisahusiwaNo ratings yet
- The Neuroscience of Tinnitus: Jos J. Eggermont and Larry E. RobertsDocument7 pagesThe Neuroscience of Tinnitus: Jos J. Eggermont and Larry E. RobertsNathalia MartinezNo ratings yet
- Batson2017 OtosclerosisDocument6 pagesBatson2017 OtosclerosisnuhajazminNo ratings yet
- Brainsci 10 00732Document11 pagesBrainsci 10 00732Nur Rahmi SeptiantyNo ratings yet
- Bhramari Pranaya and DeafnessDocument3 pagesBhramari Pranaya and DeafnesspuNo ratings yet
- Effect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing PatientsDocument6 pagesEffect of Tinnitus in Distortion Products Otoacoustic Emissions (Dpoaes) in Normal Hearing Patientsvictoria barrientoNo ratings yet
- Foto Acoustic EmitionDocument83 pagesFoto Acoustic EmitionAlok kumarNo ratings yet
- Challenges of Hearing Rehabilitation After Radiation and ChemotherapyDocument11 pagesChallenges of Hearing Rehabilitation After Radiation and ChemotherapyInes Camilla PutriNo ratings yet
- Auditory & Non Auditory Effects of Noise On Health (Mathias Barner Et. Al)Document22 pagesAuditory & Non Auditory Effects of Noise On Health (Mathias Barner Et. Al)Piyush MalviyaNo ratings yet
- Minimally Invasive Surgery For The Treatment Of.97204Document7 pagesMinimally Invasive Surgery For The Treatment Of.97204Tamilselvam KakkanaiahNo ratings yet
- AR - 09-15 - Chen 5Document10 pagesAR - 09-15 - Chen 5Arnesh kumar issarNo ratings yet
- NIH Public Access: Author ManuscriptDocument14 pagesNIH Public Access: Author ManuscriptamraovcinaNo ratings yet
- Rontal1979 Vocal Cord Dysfunction - An Industrial Health HazardDocument4 pagesRontal1979 Vocal Cord Dysfunction - An Industrial Health HazardAna BrankovićNo ratings yet
- Risk of Sound-Induced Hearing Disorders For Audio Post Production Engineers: A Preliminary StudyDocument10 pagesRisk of Sound-Induced Hearing Disorders For Audio Post Production Engineers: A Preliminary StudylauraNo ratings yet
- Recent Advancements in The Regeneration of Auditory Hair Cells and Hearing RestorationDocument16 pagesRecent Advancements in The Regeneration of Auditory Hair Cells and Hearing RestorationRana Sabouni TabariNo ratings yet
- Ultrasound EyesDocument8 pagesUltrasound Eyes-spectatorNo ratings yet
- Hearing Research: Arwa Kurabi, Elizabeth M. Keithley, Gary D. Housley, Allen F. Ryan, Ann C.-Y. WongDocument9 pagesHearing Research: Arwa Kurabi, Elizabeth M. Keithley, Gary D. Housley, Allen F. Ryan, Ann C.-Y. WongflorensiaNo ratings yet
- Noise and Its Effect On Human & Environment: Submitted ToDocument5 pagesNoise and Its Effect On Human & Environment: Submitted ToSeharAliNo ratings yet
- Safety of Ofloxacin Otic and Other Ototopical Treatments in Animal Models and in HumansDocument4 pagesSafety of Ofloxacin Otic and Other Ototopical Treatments in Animal Models and in HumansUlayya Ghina NabillaNo ratings yet
- Tinnitus and Normal Hearing: A Study On The Transient Otoacoustic Emissions SuppressionDocument6 pagesTinnitus and Normal Hearing: A Study On The Transient Otoacoustic Emissions SuppressionMarlys Ramírez ÁlvarezNo ratings yet
- Contemporary Issues in Audiology: A Hearing Scientist's PerspectiveDocument13 pagesContemporary Issues in Audiology: A Hearing Scientist's PerspectiveHarshit AmbeshNo ratings yet
- Tinnitus Heterogeneity On Auditory and Psychological Characteristics An EditorialDocument4 pagesTinnitus Heterogeneity On Auditory and Psychological Characteristics An EditorialMateus NicacioNo ratings yet
- ReversedIpsilateralAcousticReflexPattern BaslDocument9 pagesReversedIpsilateralAcousticReflexPattern BaslSarah fadlainiNo ratings yet
- Progressive Asymmetry in Occupational Noise-Induced Hearing Loss A Large Population-Based Cohort Study With A 15-Year Follow-Up PDFDocument6 pagesProgressive Asymmetry in Occupational Noise-Induced Hearing Loss A Large Population-Based Cohort Study With A 15-Year Follow-Up PDFKaty Marlen GongoraNo ratings yet
- Noise-Induced Hearing Loss in Mice: Effects of High and Low Levels of Noise Trauma in CBA MiceDocument4 pagesNoise-Induced Hearing Loss in Mice: Effects of High and Low Levels of Noise Trauma in CBA MiceJericho ChanNo ratings yet
- Spirometry 68d35cf48bffc4ef4045Document23 pagesSpirometry 68d35cf48bffc4ef4045nantousNo ratings yet
- Cromo CDCDocument592 pagesCromo CDCnantousNo ratings yet
- Fujimura2000 (00001) PDFDocument5 pagesFujimura2000 (00001) PDFnantousNo ratings yet
- The Revised Fibromyalgia Impact Questionnaire (FIQR) : Validation and Psychometric PropertiesDocument14 pagesThe Revised Fibromyalgia Impact Questionnaire (FIQR) : Validation and Psychometric PropertiesnantousNo ratings yet
- Toxicology Letters: Lists Available atDocument8 pagesToxicology Letters: Lists Available atnantousNo ratings yet
- Carol Queen - Lesbian Love in The Swingin' SeventiesDocument12 pagesCarol Queen - Lesbian Love in The Swingin' SeventiesShiri EisnerNo ratings yet
- The Central Pollution Control BoardDocument7 pagesThe Central Pollution Control BoardGanapati Joshi SankolliNo ratings yet
- Copper Alloy Guide EngDocument3 pagesCopper Alloy Guide EngRishi SharmaNo ratings yet
- EnglishDocument3 pagesEnglishšhïśh MăķwâñāNo ratings yet
- Hirschman 1982 Hedonic Consumption Emerging Concepts Methods and Propositio PDFDocument11 pagesHirschman 1982 Hedonic Consumption Emerging Concepts Methods and Propositio PDFMichael Alexandre Ramos BravoNo ratings yet
- Covid Ecri - Disinfectant Concentrations For Epa ListDocument16 pagesCovid Ecri - Disinfectant Concentrations For Epa ListNikesh ShahNo ratings yet
- Early Childhood Caries: Prevalence, Risk Factors, and PreventionDocument7 pagesEarly Childhood Caries: Prevalence, Risk Factors, and PreventioncareNo ratings yet
- CVA DVA Werkstuk Bekele - tcm39 91310Document40 pagesCVA DVA Werkstuk Bekele - tcm39 91310s_75No ratings yet
- Test Bank For Clinical Assessment in Respiratory Care 6th Edition WilkinsDocument19 pagesTest Bank For Clinical Assessment in Respiratory Care 6th Edition Wilkinsfideliamaximilian7pjjf100% (22)
- Magnetic Pulse Welding: by Gautham SarangDocument25 pagesMagnetic Pulse Welding: by Gautham SarangGauthamSarangNo ratings yet
- Agency ArchiveDocument413 pagesAgency ArchiveAZ 211100% (3)
- In Vivo Bio-Safety Evaluations and Diagnostic/Therapeutic Applications of Chemically Designed Mesoporous Silica NanoparticlesDocument33 pagesIn Vivo Bio-Safety Evaluations and Diagnostic/Therapeutic Applications of Chemically Designed Mesoporous Silica NanoparticlesAFRIZANo ratings yet
- 2011UCMC Trauma Care Q&ADocument2 pages2011UCMC Trauma Care Q&ASam CholkeNo ratings yet
- DC Generator Equation of Induced Emf (Sample Problems)Document2 pagesDC Generator Equation of Induced Emf (Sample Problems)vincent gonzalesNo ratings yet
- Lecture3 PDFDocument3 pagesLecture3 PDFKumar RajeshNo ratings yet
- YakultDocument15 pagesYakultPRIYA KUMARINo ratings yet
- Jay Dee Harp - RedactedDocument7 pagesJay Dee Harp - RedactedHeavyNo ratings yet
- Climate Change BrochureDocument2 pagesClimate Change BrochureNicole Natcher Gallego100% (1)
- T or 1155 Year 1 The Zoo Vet Differentiated Reading Comprehension Activity Ver 8Document9 pagesT or 1155 Year 1 The Zoo Vet Differentiated Reading Comprehension Activity Ver 8Dla SemecoNo ratings yet
- EmotionsDocument18 pagesEmotionsVinod DhanapalNo ratings yet
- Safety InductionDocument2 pagesSafety Inductionmark jayson yabesNo ratings yet
- Design of Columns, BeamsDocument7 pagesDesign of Columns, BeamsAbdulaashik AbdulkareemNo ratings yet
- K0821 054384 en R00 SQL-mapping-list 1CWE5336 HasanbeyliDocument11 pagesK0821 054384 en R00 SQL-mapping-list 1CWE5336 HasanbeyliRana DanialNo ratings yet
- Iridology: The Iris Can Indicate A Problem in Its Earlier Inception, Long Before Disease Symptoms Are Present.Document3 pagesIridology: The Iris Can Indicate A Problem in Its Earlier Inception, Long Before Disease Symptoms Are Present.Shoeb MohammedNo ratings yet
- K11 HS Đề Cương Ôn Tập HK1Document18 pagesK11 HS Đề Cương Ôn Tập HK1thanh_inforNo ratings yet
- Arizona Community Foundation 2011 - Arizona Organizing Project Grant ApplicationDocument5 pagesArizona Community Foundation 2011 - Arizona Organizing Project Grant ApplicationElizabeth VenableNo ratings yet
- Tutorial 1B Principal Stresses and PlanesDocument2 pagesTutorial 1B Principal Stresses and PlanesumarkamranNo ratings yet
- Visual ImpairmentDocument7 pagesVisual ImpairmentROSE MARIE REVILLA100% (1)
- PAEStigan Vol. I-XI (Review Lay-Out)Document12 pagesPAEStigan Vol. I-XI (Review Lay-Out)John Vincent Nate100% (1)
- 2023 LASC Design, Test, & Evaluation Guide R00Document32 pages2023 LASC Design, Test, & Evaluation Guide R00Juan RengifoNo ratings yet
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (4)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (39)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 4 out of 5 stars4/5 (5)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 4.5 out of 5 stars4.5/5 (83)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Rating: 3 out of 5 stars3/5 (1)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- The Twentysomething Treatment: A Revolutionary Remedy for an Uncertain AgeFrom EverandThe Twentysomething Treatment: A Revolutionary Remedy for an Uncertain AgeRating: 4.5 out of 5 stars4.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (170)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (267)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- I Shouldn't Feel This Way: Name What’s Hard, Tame Your Guilt, and Transform Self-Sabotage into Brave ActionFrom EverandI Shouldn't Feel This Way: Name What’s Hard, Tame Your Guilt, and Transform Self-Sabotage into Brave ActionNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (256)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (6)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesFrom EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesRating: 4.5 out of 5 stars4.5/5 (1412)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (44)
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (46)
- Summary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (61)