You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (346)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
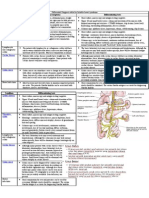
- DR Joe Chaffin Osler CourseDocument81 pagesDR Joe Chaffin Osler CourseHussein MOUSSANo ratings yet
- SyncopeDocument10 pagesSyncopeHNINNo ratings yet
- UFP Biology: Name Date of Birth CEG Number 0800 SignatureDocument10 pagesUFP Biology: Name Date of Birth CEG Number 0800 SignatureRyan JenningsNo ratings yet
- Differential DiagnosisDocument1 pageDifferential Diagnosisririz b100% (1)
- Plasma Cytokeratin 18 Level As A Novel Biomarker.5Document7 pagesPlasma Cytokeratin 18 Level As A Novel Biomarker.5HNINNo ratings yet
- Apoptosis in Hepatitis C Virus InfectionDocument11 pagesApoptosis in Hepatitis C Virus InfectionHNINNo ratings yet
- HCV EASL CPG Slide Deck 2020Document57 pagesHCV EASL CPG Slide Deck 2020HNINNo ratings yet
- Optimal Management of Anticoagulation Therapy: An Educational Slide SetDocument43 pagesOptimal Management of Anticoagulation Therapy: An Educational Slide SetHNINNo ratings yet
- Association Between Serum Cytokeratin-18 Neoepitope M30 (CK-18 M30) Levels and Chronic Hepatitis B: A Meta-AnalysisDocument7 pagesAssociation Between Serum Cytokeratin-18 Neoepitope M30 (CK-18 M30) Levels and Chronic Hepatitis B: A Meta-AnalysisHNINNo ratings yet
- WJH 11 250Document15 pagesWJH 11 250HNINNo ratings yet
- Dr. Mohammad Shaikhani. Sulaimani University, College of Medicine. Sulaimanya-Iraqi KurdistanDocument51 pagesDr. Mohammad Shaikhani. Sulaimani University, College of Medicine. Sulaimanya-Iraqi KurdistanHNINNo ratings yet
- Crabb Et Al-2020-HepatologyDocument28 pagesCrabb Et Al-2020-HepatologyHNINNo ratings yet
- Alcohol Use Disorder: Dr. Nyo Nyo Aung Senior Consultant Psychiatrist Associate Professor, YmhhDocument60 pagesAlcohol Use Disorder: Dr. Nyo Nyo Aung Senior Consultant Psychiatrist Associate Professor, YmhhHNINNo ratings yet
- Drug-Induced Liver Injury: Clinical Practice GuidelinesDocument44 pagesDrug-Induced Liver Injury: Clinical Practice GuidelinesHNINNo ratings yet
- Approach To The Cardiovascular Examination567 160120085414 PDFDocument22 pagesApproach To The Cardiovascular Examination567 160120085414 PDFHNINNo ratings yet
- Document PDFDocument273 pagesDocument PDFHNINNo ratings yet
- Hepatology: World Journal ofDocument18 pagesHepatology: World Journal ofHNINNo ratings yet
- Hepatitis B and C Virus-Induced Hepatitis: Apoptosis, Autophagy, and Unfolded Protein ResponseDocument16 pagesHepatitis B and C Virus-Induced Hepatitis: Apoptosis, Autophagy, and Unfolded Protein ResponseHNINNo ratings yet
- WJH-11-421 2Document25 pagesWJH-11-421 2HNINNo ratings yet
- Sample 8082Document11 pagesSample 8082Devil SinghNo ratings yet
- 21 Health Benefits of Bitter KolaDocument4 pages21 Health Benefits of Bitter Kolaserjio47No ratings yet
- Nonsteroidal Anti-Inflamatory DrugsDocument4 pagesNonsteroidal Anti-Inflamatory DrugsYogi drNo ratings yet
- Openstax College Concepts of Biology Supplemental Test Items Chapter 09: Molecular BiologyDocument4 pagesOpenstax College Concepts of Biology Supplemental Test Items Chapter 09: Molecular BiologyFatima BjaijihNo ratings yet
- Serum Protein ElectrophoresisDocument9 pagesSerum Protein Electrophoresiskiedd_04100% (4)
- Reviewer in PhCh2Document9 pagesReviewer in PhCh2prinz1mendezNo ratings yet
- Genetics Learning Materials Bsa 2 1Document34 pagesGenetics Learning Materials Bsa 2 1Khy Renz Bajaj100% (1)
- Learning Unit 2 Prokaryotic Genome SystemDocument22 pagesLearning Unit 2 Prokaryotic Genome SystemgengkapakNo ratings yet
- 1.2 Nucleic Acids, Biology For Engineers DRPDocument4 pages1.2 Nucleic Acids, Biology For Engineers DRPShreya shresthNo ratings yet
- Acute Lymphoblastic Leukemia Characterizationandits Prognostic ValuesDocument11 pagesAcute Lymphoblastic Leukemia Characterizationandits Prognostic ValuesDebby ApriliaNo ratings yet
- Proceeding of The NAVC North American Veterinary Conference: Jan. 13-27, 2007, Orlando, FloridaDocument4 pagesProceeding of The NAVC North American Veterinary Conference: Jan. 13-27, 2007, Orlando, FloridaLuis SanchezNo ratings yet
- May Kulang Pa Sa METHODOLOGY, RESULTS, DISCUSSION, RECOMMENDATIONDocument16 pagesMay Kulang Pa Sa METHODOLOGY, RESULTS, DISCUSSION, RECOMMENDATIONAnna Garcia BuquidNo ratings yet
- Zinc Supplementation and COVID-19: RecommendationsDocument4 pagesZinc Supplementation and COVID-19: RecommendationsAdrian CalinNo ratings yet
- Chapter 14 - in Silico Identification of Small RNAs A - 2022 - Bioinformatics IDocument9 pagesChapter 14 - in Silico Identification of Small RNAs A - 2022 - Bioinformatics IMuhammad KamranNo ratings yet
- Coronaviruses Methods and Protocols - Helena Jane Maier 2015Document299 pagesCoronaviruses Methods and Protocols - Helena Jane Maier 2015Geronimo Sanchez VicarioNo ratings yet
- Journal Pre-Proof: European Journal of Medicinal ChemistryDocument70 pagesJournal Pre-Proof: European Journal of Medicinal ChemistryKristina MilićevićNo ratings yet
- MutagenDocument15 pagesMutagenDibyakNo ratings yet
- Study Guide For Lecture Exam 2 Biol 1407 03.12.2010Document50 pagesStudy Guide For Lecture Exam 2 Biol 1407 03.12.2010Erica MizzIndependent BubuchuNo ratings yet
- Nursing2021 Drug Handbook®, 41eDocument73 pagesNursing2021 Drug Handbook®, 41eangel marycor valdezNo ratings yet
- Building Better Monoclonal Antibody-Based Therapeutics: George J. WeinerDocument10 pagesBuilding Better Monoclonal Antibody-Based Therapeutics: George J. WeinerSyahputraWibowoNo ratings yet
- Injoere v2n2p46 enDocument33 pagesInjoere v2n2p46 enAndia BalabanNo ratings yet
- UNIT 5 Students Topic 1 and 2 NotesDocument25 pagesUNIT 5 Students Topic 1 and 2 Noteskayecec996No ratings yet
- Activity 7 - Cell Cycle & Cell Division PDFDocument3 pagesActivity 7 - Cell Cycle & Cell Division PDFKiro ZeroNo ratings yet
- 2019Document53 pages2019Hey Its Coby0% (1)
- Cie155-2003 (Free Copy March 2020) PDFDocument74 pagesCie155-2003 (Free Copy March 2020) PDFmiro55slavNo ratings yet
- Subcutaneous MycosesDocument17 pagesSubcutaneous MycosesSit IsahNo ratings yet
- CBSE Class 12 Biology Chapter 8 Human Health and Disease Revision NotesDocument48 pagesCBSE Class 12 Biology Chapter 8 Human Health and Disease Revision Notesdevesh saiNo ratings yet