You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5811)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Full Ebook of Vertebrate Life 11Th Edition F Harvey Pough Online PDF All ChapterDocument69 pagesFull Ebook of Vertebrate Life 11Th Edition F Harvey Pough Online PDF All Chapterfawakarelex100% (10)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Invertebrates Brusca (2018-Ingles) PDFDocument1,128 pagesInvertebrates Brusca (2018-Ingles) PDFSarah Lucia Arroyave Moreno89% (9)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Class #1 Eng III G 23Document19 pagesClass #1 Eng III G 23RODRIGUEZ RODRIGUEZ MICHAEL STIVENNo ratings yet
- A Multi-Model Framework For Climate Change Impact AssessmentDocument16 pagesA Multi-Model Framework For Climate Change Impact AssessmentRODRIGUEZ RODRIGUEZ MICHAEL STIVENNo ratings yet
- Adaptation According To Mode of Climate Variability: A Case Study From Canada 'S Western InteriorDocument24 pagesAdaptation According To Mode of Climate Variability: A Case Study From Canada 'S Western InteriorRODRIGUEZ RODRIGUEZ MICHAEL STIVENNo ratings yet
- Accepted ManuscriptDocument38 pagesAccepted ManuscriptRODRIGUEZ RODRIGUEZ MICHAEL STIVENNo ratings yet
- Barito Kutai Basin Stratigraphic ColumnDocument1 pageBarito Kutai Basin Stratigraphic ColumnAkarius MagersNo ratings yet
- Trex TUTORIALstencilDocument4 pagesTrex TUTORIALstencilSandra ValeNo ratings yet
- Cerda Et Al - 2021Document17 pagesCerda Et Al - 2021Jose L CarballidoNo ratings yet
- Mammals From The Age of Dinosaurs OriginDocument2 pagesMammals From The Age of Dinosaurs OriginLili BujorNo ratings yet
- Classification and Evolution of Living Things 2Document100 pagesClassification and Evolution of Living Things 2axelle89017No ratings yet
- Concepts of Biology 3rd Edition Mader Solutions Manual 1Document2 pagesConcepts of Biology 3rd Edition Mader Solutions Manual 1barbara100% (36)
- Ms. Susan M. Pojer Horace Greeley HS Chappaqua, NYDocument30 pagesMs. Susan M. Pojer Horace Greeley HS Chappaqua, NYII-2 Saint ClareNo ratings yet
- Chemical Mystery 1Document3 pagesChemical Mystery 1Keirby CayanongNo ratings yet
- The "Tully Monster" ReconstructionDocument12 pagesThe "Tully Monster" ReconstructionAndrew KirkNo ratings yet
- Mohammad Ashraf - 2010 - Petrographic and Biostratigraphic Studies On Sedimentary Sequences of Multanai Area, Pishin BasinDocument12 pagesMohammad Ashraf - 2010 - Petrographic and Biostratigraphic Studies On Sedimentary Sequences of Multanai Area, Pishin BasinShakeel MirwaniNo ratings yet
- Pendidikan Matematika - Unit 7 - Group East Lombok 1Document3 pagesPendidikan Matematika - Unit 7 - Group East Lombok 1Muhammad Kasyiful KurabiNo ratings yet
- Ade Rizky - 270110190138 - E - Tugas 3 Dan 4Document2 pagesAde Rizky - 270110190138 - E - Tugas 3 Dan 4Ade RizkyNo ratings yet
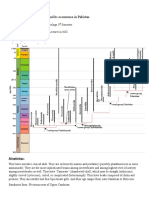
- Cephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanDocument2 pagesCephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanEEsha AhmedNo ratings yet
- Geologic Time - Webquest 2020Document3 pagesGeologic Time - Webquest 2020Patrick SazamaNo ratings yet
- Human Origins by Alan Walker and Pat Shipman, The Main Focus Was On How Brains and Diet AreDocument1 pageHuman Origins by Alan Walker and Pat Shipman, The Main Focus Was On How Brains and Diet Areirc32393No ratings yet
- A New Species of Pachyrhinosaurus CeratoDocument254 pagesA New Species of Pachyrhinosaurus CeratoJose Perez LopezNo ratings yet
- Vergara - Et - Al - 1995 - Bulletin - GSADocument14 pagesVergara - Et - Al - 1995 - Bulletin - GSAEngell CavadaNo ratings yet
- Detailed Lesson Plan UCSP WEEK 2 ContinuationDocument15 pagesDetailed Lesson Plan UCSP WEEK 2 Continuationchris wangNo ratings yet
- ExtinctionDocument5 pagesExtinctionKhairul ShahmiNo ratings yet
- Alam Et Al 2003Document30 pagesAlam Et Al 2003Furious SKNo ratings yet
- Dredged Up 28, Spring 2021Document8 pagesDredged Up 28, Spring 2021Wessex ArchaeologyNo ratings yet
- Eons - BiologyDocument4 pagesEons - BiologyA - UNO, France Jhondale B.No ratings yet
- Liuetal HaootiaDocument11 pagesLiuetal HaootiaFabio FotografNo ratings yet
- Power Functions - ClasszoneDocument6 pagesPower Functions - ClasszoneAnoop SreedharNo ratings yet
- Quiz MakingDocument9 pagesQuiz MakingKyle RaNo ratings yet
- Bogicevic Et Al. - 2016 - Preliminary Results of The Analysis of PleistoceneDocument6 pagesBogicevic Et Al. - 2016 - Preliminary Results of The Analysis of Pleistocenehommie pearNo ratings yet
- The CephalopodaDocument8 pagesThe CephalopodaAtifAliBukhariNo ratings yet
- Al Jahagbub Paper, 2022Document24 pagesAl Jahagbub Paper, 2022محمد الحاسيNo ratings yet