You might also like
- Euroscience of Usic and Motion: Gunter Kreutz and Martin LotzeDocument25 pagesEuroscience of Usic and Motion: Gunter Kreutz and Martin LotzeRandall GloverNo ratings yet
- Juslin - Emotional Responses To MusicDocument49 pagesJuslin - Emotional Responses To MusicVeruschka MainhardNo ratings yet
- Juslin Ve Diğ., 2014Document25 pagesJuslin Ve Diğ., 2014Meriç EsenNo ratings yet
- Psychophysiological Responses To Happy and Sad MusicDocument16 pagesPsychophysiological Responses To Happy and Sad MusicGonzalo Alonso OsmaNo ratings yet
- What Makes Music Emotionally SignificantDocument26 pagesWhat Makes Music Emotionally SignificantEduardo Mello0% (1)
- Music Emotional Responses ToDocument63 pagesMusic Emotional Responses ToJunior Leme100% (1)
- Juslin BehavBrSci31 2008 Emotional-Reponses-To-MusicDocument64 pagesJuslin BehavBrSci31 2008 Emotional-Reponses-To-MusicMauricio De BaileNo ratings yet
- Juslin y Laukka (2004) - Expression Perception and Induction of Musical Emotions A Review and A Questionnaire Study of Everyday ListeningDocument23 pagesJuslin y Laukka (2004) - Expression Perception and Induction of Musical Emotions A Review and A Questionnaire Study of Everyday ListeningMaría MarchianoNo ratings yet
- Livro Juslin e Sloboda - Psicologia Da MusicaDocument8 pagesLivro Juslin e Sloboda - Psicologia Da MusicaMEIRE HADADNo ratings yet
- Two LevelModelofEmbodidCognitioninMuscKorsakova Kreyn2018Document49 pagesTwo LevelModelofEmbodidCognitioninMuscKorsakova Kreyn2018Alicia MenesesNo ratings yet
- Juslin, P. N., & Vastfjall, D. (2008) - Emotional Responses To Music - The Need To Consider Underlying Mechanisms. Behavioral and Brain Sciences, 31 (5), 559.Document63 pagesJuslin, P. N., & Vastfjall, D. (2008) - Emotional Responses To Music - The Need To Consider Underlying Mechanisms. Behavioral and Brain Sciences, 31 (5), 559.goni56509No ratings yet
- Daltrozzo Et Al 2011Document14 pagesDaltrozzo Et Al 2011RaminIsmailoffNo ratings yet
- A Neurodynamic Perspective On Musical en PDFDocument6 pagesA Neurodynamic Perspective On Musical en PDFHeoel GreopNo ratings yet
- Reflections On Music Affect and SocialityDocument20 pagesReflections On Music Affect and SocialityruiafaNo ratings yet
- 2b Enacting-Musical-EmotionsDocument25 pages2b Enacting-Musical-EmotionsYuri Alexndra Alfonso CastiblancoNo ratings yet
- Music AnalysisDocument13 pagesMusic AnalysisbicibassNo ratings yet
- 0b Music and Its Inductive Power - A Psychobiological and Evolutionary Approach To Musical EmotionsDocument15 pages0b Music and Its Inductive Power - A Psychobiological and Evolutionary Approach To Musical EmotionsLuis Felipe Rodriguez CortesNo ratings yet
- Mapping Aesthetic Musical Emotions in The BrainDocument15 pagesMapping Aesthetic Musical Emotions in The BrainFrancesco CiongoliNo ratings yet
- Music and Emotions ThesisDocument5 pagesMusic and Emotions Thesisbkx89b3r100% (2)
- Neural Mechanisms of Music TherapyDocument14 pagesNeural Mechanisms of Music TherapyMae P100% (1)
- The Pleasure of Reading: Morten L. Kringelbach, Peter Vuust and John GeakeDocument15 pagesThe Pleasure of Reading: Morten L. Kringelbach, Peter Vuust and John GeakeAxel JuárezNo ratings yet
- Zentner, Grandjean & Scherer (2008) Emotions Evoked by The Sound of MusicDocument28 pagesZentner, Grandjean & Scherer (2008) Emotions Evoked by The Sound of MusicregimarcosNo ratings yet
- Emotional Responses To Music: Experience, Expression, and PhysiologyDocument30 pagesEmotional Responses To Music: Experience, Expression, and PhysiologyArooba fatimaNo ratings yet
- 125982068Document6 pages125982068godoflightningzenitsuNo ratings yet
- Emotional Responses To Music: Experience, Expression, and PhysiologyDocument30 pagesEmotional Responses To Music: Experience, Expression, and PhysiologyVishal SeksariaNo ratings yet
- Tunning The Brain For MusicDocument3 pagesTunning The Brain For Musicjohn fredy perez100% (1)
- Perceptual Analysis MozartDocument33 pagesPerceptual Analysis MozartnicholasNo ratings yet
- 10-1108 03055720810904817人类知识系统:音乐与大脑连贯性Document19 pages10-1108 03055720810904817人类知识系统:音乐与大脑连贯性陈宝利No ratings yet
- Emotional Induction Through MusicDocument23 pagesEmotional Induction Through MusicZane JoinerNo ratings yet
- Towards A Neural Basis of Music-Evoked EmotionsDocument7 pagesTowards A Neural Basis of Music-Evoked EmotionsKevin Stephenson100% (1)
- Exploring Emotional Reactions to Music MechanismsDocument25 pagesExploring Emotional Reactions to Music MechanismsCrhistian Esteban Hidalgo ValbuenaNo ratings yet
- A Composition Informed by A Study On The Emotional Power of Music - Dimitrios MylisDocument131 pagesA Composition Informed by A Study On The Emotional Power of Music - Dimitrios MylisDimitris MylisNo ratings yet
- Brain and The MusicDocument28 pagesBrain and The MusicLuzia liquerNo ratings yet
- Basic EmotionDocument9 pagesBasic EmotionRakib RabiNo ratings yet
- Emotions Evoked by The Sound of MusicDocument29 pagesEmotions Evoked by The Sound of MusicΕιρηνη ΚαλογερακηNo ratings yet
- The Future of Musical EmotionsDocument5 pagesThe Future of Musical EmotionsRael Gimenes ToffoloNo ratings yet
- The Psychology of Entertainment Media Blurring The Lines Between Entertainment and PersuasionDocument32 pagesThe Psychology of Entertainment Media Blurring The Lines Between Entertainment and PersuasionJoeNo ratings yet
- Zatorre, R. J. (2003) - Music and The Brain. Annals of The New York Academy of Sciences, 999 (1), 4-14.Document11 pagesZatorre, R. J. (2003) - Music and The Brain. Annals of The New York Academy of Sciences, 999 (1), 4-14.goni56509No ratings yet
- Groove Experience: Emotional and Physiological Responses To Groove-Based MusicDocument10 pagesGroove Experience: Emotional and Physiological Responses To Groove-Based MusicBruna Siqueira FrançaNo ratings yet
- Research Paper 1Document18 pagesResearch Paper 1api-515601372No ratings yet
- Neurologic Music Therapy in Cognitive RehabilitationDocument7 pagesNeurologic Music Therapy in Cognitive Rehabilitationisaacelias2509No ratings yet
- Embodied cognition in music explained through tonal relationships and tensionDocument14 pagesEmbodied cognition in music explained through tonal relationships and tensionAlicia Meneses100% (1)
- Music Cognition Research ApproachesDocument52 pagesMusic Cognition Research ApproachesRaul Alejandro Celis MaturanaNo ratings yet
- The Psychological Functions of Music Listening: Thomas Schäfer, Peter Sedlmeier, Christine Städtler and David HuronDocument33 pagesThe Psychological Functions of Music Listening: Thomas Schäfer, Peter Sedlmeier, Christine Städtler and David Huronaavriti6kushwahaNo ratings yet
- Mapping Musical Aesthetic Emotion in The BrainDocument15 pagesMapping Musical Aesthetic Emotion in The BrainNaji EssmaeelNo ratings yet
- Eduardo Lopes Time Rhythm and Meter in Music The Gravitational ConceptDocument14 pagesEduardo Lopes Time Rhythm and Meter in Music The Gravitational Conceptjrhee88No ratings yet
- University of California Press Music Perception: An Interdisciplinary JournalDocument19 pagesUniversity of California Press Music Perception: An Interdisciplinary JournalArnold MukaratiNo ratings yet
- Brain-Music Connection: Activations That Continue To Amaze: Recurring MotifsDocument3 pagesBrain-Music Connection: Activations That Continue To Amaze: Recurring MotifsAli AvşarNo ratings yet
- Gale Researcher Guide for: Experimental and Behavioral PsychologyFrom EverandGale Researcher Guide for: Experimental and Behavioral PsychologyNo ratings yet
- Music, Memory and EmotionDocument6 pagesMusic, Memory and EmotionLuciane CuervoNo ratings yet
- Investigating Relationships Between Music, Emotions, Personality, and Music-Induced MovementDocument11 pagesInvestigating Relationships Between Music, Emotions, Personality, and Music-Induced MovementRenata RatnasariNo ratings yet
- Patel - 2010 - Music, Biological Evolution, and The BrainDocument37 pagesPatel - 2010 - Music, Biological Evolution, and The BrainMauricio SanchezNo ratings yet
- Neural Correlates of Music Listening: Does The Music Matter?Document20 pagesNeural Correlates of Music Listening: Does The Music Matter?Mark ReybrouckNo ratings yet
- Author's Accepted Manuscript: NeuropsychologiaDocument50 pagesAuthor's Accepted Manuscript: NeuropsychologiaPaolo BertoliniNo ratings yet
- Music and the brain_ the neuroscience of music and musical appreciation - PMCDocument5 pagesMusic and the brain_ the neuroscience of music and musical appreciation - PMCAlexNo ratings yet
- Emotion and Delusion: Seeking Common Ground Between Neuroscience and The PsychotherapiesDocument22 pagesEmotion and Delusion: Seeking Common Ground Between Neuroscience and The PsychotherapiesVeronica CaselliNo ratings yet
- Neuroscience of music perception and productionDocument10 pagesNeuroscience of music perception and productionKeven GaoNo ratings yet
- Daniel Västfjäll - Emotion Induction Through Music - A Review of The Musical Mood Induction ProcedureDocument48 pagesDaniel Västfjäll - Emotion Induction Through Music - A Review of The Musical Mood Induction ProcedureFrancisVincentNo ratings yet
- ERQ Study Manuscript ACCEPTEDDocument30 pagesERQ Study Manuscript ACCEPTEDmarcelocarvalhoteacher2373No ratings yet
- Uusberg Uusberg GrossDocument25 pagesUusberg Uusberg Grossmarcelocarvalhoteacher2373No ratings yet
- Validacion ErqDocument7 pagesValidacion ErqMariaNo ratings yet
- Neurobiology of Fear and Specific PhobiasDocument10 pagesNeurobiology of Fear and Specific Phobiasmarcelocarvalhoteacher2373No ratings yet
- Organizational Change and Employee Stress PDFDocument41 pagesOrganizational Change and Employee Stress PDFmarcelocarvalhoteacher2373No ratings yet
- Modulation of Human Natural Killer Cell Activity by Exposure To Uncontrollable Stress'Document16 pagesModulation of Human Natural Killer Cell Activity by Exposure To Uncontrollable Stress'marcelocarvalhoteacher2373No ratings yet
- More Than A Feeling A Unified View of ST PDFDocument24 pagesMore Than A Feeling A Unified View of ST PDFmarcelocarvalhoteacher2373No ratings yet
- Neurobiology of Fear and Specific PhobiasDocument10 pagesNeurobiology of Fear and Specific Phobiasmarcelocarvalhoteacher2373No ratings yet
- Unrealistic Optimism About Future Life Events A CaDocument21 pagesUnrealistic Optimism About Future Life Events A Camarcelocarvalhoteacher2373No ratings yet
- Eustress Vs Distress-A ReviewDocument4 pagesEustress Vs Distress-A Reviewmarcelocarvalhoteacher2373No ratings yet
- The Measurement of Optimism and Hope in Relation To College StudeDocument91 pagesThe Measurement of Optimism and Hope in Relation To College StudeRasmi Ranjan MuduliNo ratings yet
- Eustress Distress Appraisal Scale PDFDocument13 pagesEustress Distress Appraisal Scale PDFmarcelocarvalhoteacher2373No ratings yet
- Journal of Business Research: SciencedirectDocument7 pagesJournal of Business Research: Sciencedirectmarcelocarvalhoteacher2373No ratings yet
- Positive Psychotherapy: American Psychologist January 2016Document16 pagesPositive Psychotherapy: American Psychologist January 2016marcelocarvalhoteacher2373No ratings yet
- Optimism, Goal Conflict, and Stressor-Related Immune Change: Suzanne C. SegerstromDocument27 pagesOptimism, Goal Conflict, and Stressor-Related Immune Change: Suzanne C. Segerstrommarcelocarvalhoteacher2373No ratings yet
- Measuring Optimism in the WorkplaceDocument23 pagesMeasuring Optimism in the Workplacemarcelocarvalhoteacher2373No ratings yet
- Severe Covid-19: Clinical PracticeDocument10 pagesSevere Covid-19: Clinical Practicemarcelocarvalhoteacher2373No ratings yet
- A Positive Approach To Stress PDFDocument5 pagesA Positive Approach To Stress PDFmarcelocarvalhoteacher2373No ratings yet
- Current Issues in Uncertainty PDFDocument6 pagesCurrent Issues in Uncertainty PDFmarcelocarvalhoteacher2373No ratings yet
- Xerostomia and Its Management: S.AbhiramDocument27 pagesXerostomia and Its Management: S.Abhirammarcelocarvalhoteacher2373No ratings yet
- 2006 Arawak Languages Encyclopedia of Language and LinguisticsDocument4 pages2006 Arawak Languages Encyclopedia of Language and LinguisticsSheldon JungleNo ratings yet
- MEDHERB - Volume 8 - Issue 2 - Pages 71-78Document8 pagesMEDHERB - Volume 8 - Issue 2 - Pages 71-78Andreea RoxanaNo ratings yet
- How Did Sundar Pichai Become CEO of Google 13102020 113057amDocument4 pagesHow Did Sundar Pichai Become CEO of Google 13102020 113057amMaherban HaiderNo ratings yet
- Analyzing Your Game Performance Using Event Tracing For WindowsDocument54 pagesAnalyzing Your Game Performance Using Event Tracing For WindowsWahyu ShanNo ratings yet
- Oracle 1z0-986 Exam: Implementation EssentialsDocument23 pagesOracle 1z0-986 Exam: Implementation Essentialslakshmanan84No ratings yet
- Selection of Hazard Evaluation Techniques PDFDocument16 pagesSelection of Hazard Evaluation Techniques PDFdediodedNo ratings yet
- Classroom Management and DisciplineDocument2 pagesClassroom Management and DisciplineMonaida Umpar IbrahimNo ratings yet
- Dr. Kirti MatliwalaDocument6 pagesDr. Kirti MatliwalaKirti Dakshesh ThakarNo ratings yet
- BS en 01062-3-2008 PDFDocument12 pagesBS en 01062-3-2008 PDFjohnNo ratings yet
- TM55 1520 242 MTFDocument192 pagesTM55 1520 242 MTFJakub Kostka100% (1)
- United States Patent (19) : (52) U.S. C.260/583 KDocument5 pagesUnited States Patent (19) : (52) U.S. C.260/583 KkurniawanNo ratings yet
- Health Teaching Plan - AttitudeDocument2 pagesHealth Teaching Plan - AttitudeFaller TrixieNo ratings yet
- LF ReportDocument32 pagesLF ReportKrisLarrNo ratings yet
- Evangelion Cruel Angels Thesis - Moonbase AlphaDocument5 pagesEvangelion Cruel Angels Thesis - Moonbase Alphasionothewil1977100% (1)
- 1 Calimutan v. People - G.R. No. 152133Document1 page1 Calimutan v. People - G.R. No. 152133eiram23100% (1)
- SAP FICO Resume - Exp - 3.5 YearsDocument3 pagesSAP FICO Resume - Exp - 3.5 Yearsranga67% (3)
- Acute Ankle Sprain - An UpdateDocument7 pagesAcute Ankle Sprain - An UpdateFran Leiva CorreaNo ratings yet
- Ich Validation 2qaDocument9 pagesIch Validation 2qaBishal AdhikariNo ratings yet
- The Role of Social Media On Leisure Preferences - A Research On The Participants of Outdoor Recreation Activities (#263492) - 230009Document10 pagesThe Role of Social Media On Leisure Preferences - A Research On The Participants of Outdoor Recreation Activities (#263492) - 230009Stella PadillaNo ratings yet
- Canon Sus 2016 e 03Document2 pagesCanon Sus 2016 e 03Roxanne Tacy SantiagoNo ratings yet
- Ambien Withdrawal TreatmentDocument6 pagesAmbien Withdrawal TreatmentSurvival SabariNo ratings yet
- Official ResumeDocument2 pagesOfficial Resumeapi-385631007No ratings yet
- List of US 501 (C) (3) Charities Located in CanadaDocument5 pagesList of US 501 (C) (3) Charities Located in CanadaART'S PLACENo ratings yet
- Pasyon and RevolutionDocument2 pagesPasyon and RevolutionBilly JoeNo ratings yet
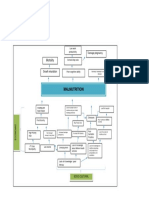
- Problem Tree Analysis MalnutritionDocument1 pageProblem Tree Analysis MalnutritionMho Dsb50% (6)
- Response To 'Enforced Sunday Law Will Soon Become The Mark of The Beast'Document22 pagesResponse To 'Enforced Sunday Law Will Soon Become The Mark of The Beast'jennywren3177No ratings yet
- The Snouters - Form and Life of - Harald StumpkeDocument126 pagesThe Snouters - Form and Life of - Harald StumpkeAlberto Humanes MuñozNo ratings yet
- 3rd Quarter Las in Science 5Document4 pages3rd Quarter Las in Science 5Michael Edward De VillaNo ratings yet
- Explicitation and Implicitation - Omission and Addition - in ENGLISHDocument63 pagesExplicitation and Implicitation - Omission and Addition - in ENGLISHdyna bookNo ratings yet
- Mark Scheme (Results) : Summer 2018Document22 pagesMark Scheme (Results) : Summer 2018RafaNo ratings yet