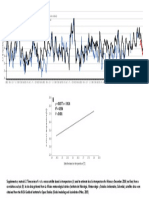
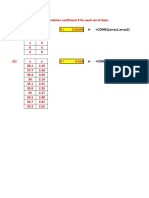
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5807)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Midterm Fall2011Document13 pagesMidterm Fall2011Patrick BensonNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 9.1 Significance Tests: The Basics: Problem 1 - 911 CallsDocument15 pages9.1 Significance Tests: The Basics: Problem 1 - 911 Callsfire historyNo ratings yet
- Hypothesis TestingDocument59 pagesHypothesis TestingHans Sicangco63% (24)
- STAT100 Fall19 Test 2 ANSWERS Practice Problems PDFDocument23 pagesSTAT100 Fall19 Test 2 ANSWERS Practice Problems PDFabutiNo ratings yet
- Fmars 08 670354Document15 pagesFmars 08 670354cr rNo ratings yet
- Mathematical Modelling of Spatial-Ecological Complex Systems: An EvaluationDocument14 pagesMathematical Modelling of Spatial-Ecological Complex Systems: An Evaluationcr rNo ratings yet
- What Are The Roles of Species Distribution Models in Conservation PlanningDocument5 pagesWhat Are The Roles of Species Distribution Models in Conservation Planningcr rNo ratings yet
- Species Distribution Models: Ecological Explanation and Prediction Across Space and TimeDocument24 pagesSpecies Distribution Models: Ecological Explanation and Prediction Across Space and Timecr rNo ratings yet
- Improving Models of Species Ecological Niches A Remote Sensing OverviewDocument7 pagesImproving Models of Species Ecological Niches A Remote Sensing Overviewcr rNo ratings yet
- Global Ecology and Conservation: Yarelys Ferrer-Sánchez, Ricardo Rodríguez-EstrellaDocument12 pagesGlobal Ecology and Conservation: Yarelys Ferrer-Sánchez, Ricardo Rodríguez-Estrellacr rNo ratings yet
- 1 s2.0 S0304377017303303 mmc2Document1 page1 s2.0 S0304377017303303 mmc2cr rNo ratings yet
- Mashimoetal 2014advanceDocument24 pagesMashimoetal 2014advancecr rNo ratings yet
- Alberti - 2020 - The Complexity of Urban Eco-Evolutionary DynamicsDocument22 pagesAlberti - 2020 - The Complexity of Urban Eco-Evolutionary Dynamicscr rNo ratings yet
- Dallai Mating Zorotypus2013Document15 pagesDallai Mating Zorotypus2013cr rNo ratings yet
- Transgene Biocontainment Strategies For Molecular FarmingDocument11 pagesTransgene Biocontainment Strategies For Molecular Farmingcr rNo ratings yet
- Escenarios NormativosDocument14 pagesEscenarios Normativoscr rNo ratings yet
- Global Trends in Mangrove Forest FragmentationDocument8 pagesGlobal Trends in Mangrove Forest Fragmentationcr rNo ratings yet
- A World Without MangrovesDocument4 pagesA World Without Mangrovescr rNo ratings yet
- A Retrospective Analysis of Mangrove Extent Estimations in Colombian Pacific MangrovesDocument1 pageA Retrospective Analysis of Mangrove Extent Estimations in Colombian Pacific Mangrovescr rNo ratings yet
- Estuarine, Coastal and Shelf Science: A A B A C D eDocument21 pagesEstuarine, Coastal and Shelf Science: A A B A C D ecr rNo ratings yet
- Remotesensing 11 02697Document20 pagesRemotesensing 11 02697cr rNo ratings yet
- Remote SensingDocument27 pagesRemote Sensingcr rNo ratings yet
- British Journal of Marketing Studies Vol.2, No.6, pp.52-67, October 2014 Published by European Centre For Research Training and Development UKDocument16 pagesBritish Journal of Marketing Studies Vol.2, No.6, pp.52-67, October 2014 Published by European Centre For Research Training and Development UKAbay ShiferaNo ratings yet
- 1) Intro To MinitabDocument29 pages1) Intro To MinitabVinayNo ratings yet
- 2022 SUMATH3020 Day 18Document4 pages2022 SUMATH3020 Day 18Abby HorneNo ratings yet
- A Doe/Qbd Optimization Model of "Hard Gelatin Capsule Encapsulation" Process Using Box Behnken RSM For Development of Hard Gelatin CapsuleDocument11 pagesA Doe/Qbd Optimization Model of "Hard Gelatin Capsule Encapsulation" Process Using Box Behnken RSM For Development of Hard Gelatin CapsuleMartin CelestinoNo ratings yet
- A Study On Financial Planning For Salaried EmployeesDocument8 pagesA Study On Financial Planning For Salaried EmployeesHeena GuptaNo ratings yet
- The T-Test For Correlated SamplesDocument9 pagesThe T-Test For Correlated SamplesSeranicoustic BandNo ratings yet
- Abiolaadeniyi 2009Document7 pagesAbiolaadeniyi 2009mutiara hapkaNo ratings yet
- Chemistry: Council For The Indian School Certificate ExaminationsDocument42 pagesChemistry: Council For The Indian School Certificate ExaminationsRudra SahaNo ratings yet
- QuestionnaireDocument72 pagesQuestionnaireSatishKakarlaNo ratings yet
- STAT2201 - Ch. 14 Excel Files - UpdatedDocument41 pagesSTAT2201 - Ch. 14 Excel Files - UpdatedGurjot SinghNo ratings yet
- Jayathilaka (2020)Document10 pagesJayathilaka (2020)Luis Jhonny Dávila ValderaNo ratings yet
- q3 Weeks56 III Learning MaterialDocument7 pagesq3 Weeks56 III Learning MaterialJohn Christopher JaroNo ratings yet
- Fisher's Exact TestDocument9 pagesFisher's Exact TestShahzad Asghar ArainNo ratings yet
- Molasses As Retarding Admixture For Concrete in Road Construction PDFDocument11 pagesMolasses As Retarding Admixture For Concrete in Road Construction PDFSha SibayanNo ratings yet
- Product Differentiation PDFDocument24 pagesProduct Differentiation PDFNancy OnuigboNo ratings yet
- Sewing Room Defects PDFDocument6 pagesSewing Room Defects PDFAlemu HaileNo ratings yet
- Analysis On Pre Merger and Post Merger Financial Performance of Selected Banks in IndiaDocument57 pagesAnalysis On Pre Merger and Post Merger Financial Performance of Selected Banks in IndiaSimran TalrejaNo ratings yet
- A Study On Milk Producing Farmer's Perception Towards Komul With Special Reference To Kolardist, KarnatakaDocument12 pagesA Study On Milk Producing Farmer's Perception Towards Komul With Special Reference To Kolardist, KarnatakaNagarjuna JNo ratings yet
- Apa AnovaDocument2 pagesApa AnovaSana JuttNo ratings yet
- RDC - Thesis and Dissertation IMRAD Format 1Document151 pagesRDC - Thesis and Dissertation IMRAD Format 1DENVER ALIGANo ratings yet
- STAT2001 HomeWork#2 ActualDocument2 pagesSTAT2001 HomeWork#2 ActualKenya LevyNo ratings yet
- A Tutorial On Conducting and Interpreting A Bayesian ANOVA in JASPDocument25 pagesA Tutorial On Conducting and Interpreting A Bayesian ANOVA in JASPAniela VizitiuNo ratings yet
- Research Report Perception Towards MBADocument41 pagesResearch Report Perception Towards MBABikram Prajapati100% (3)
- Supplementary Modules in Grammar Through Literature As Integrated Lessons For Grade 7 EnglishDocument12 pagesSupplementary Modules in Grammar Through Literature As Integrated Lessons For Grade 7 EnglishPsychology and Education: A Multidisciplinary JournalNo ratings yet
- Entrepreneurial Orientation Competitive AdvantageDocument16 pagesEntrepreneurial Orientation Competitive AdvantageAch Firman WahyudiNo ratings yet
- 16 Articulo SensorialDocument11 pages16 Articulo SensorialValeeMendezPadillaNo ratings yet