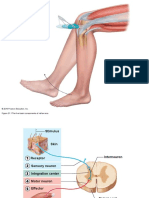
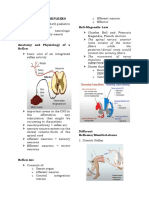
You might also like
- EndExMaleLibido PDFDocument26 pagesEndExMaleLibido PDFAdam RahimiNo ratings yet
- J Appl Physiol-2008-Kjær-1026-7Document3 pagesJ Appl Physiol-2008-Kjær-1026-7Luciana AlécioNo ratings yet
- Estrogen Effects on Traumatic Brain Injury: Mechanisms of Neuroprotection and RepairFrom EverandEstrogen Effects on Traumatic Brain Injury: Mechanisms of Neuroprotection and RepairKelli A DuncanNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptFrontiersNo ratings yet
- Sex Differences in PhysiologyFrom EverandSex Differences in PhysiologyGretchen NeighNo ratings yet
- Voss Et Al. 2011 Exercise Brain and Cognition Across The Life SpanDocument10 pagesVoss Et Al. 2011 Exercise Brain and Cognition Across The Life SpanHenriqueNo ratings yet
- Exercise, Brain, and Cognition Across The Life SpanDocument9 pagesExercise, Brain, and Cognition Across The Life SpanFernandaNo ratings yet
- Myths and Methodologies: Reducing Scientific Design Ambiguity in Studies Comparing Sexes And/or Menstrual Cycle PhasesDocument9 pagesMyths and Methodologies: Reducing Scientific Design Ambiguity in Studies Comparing Sexes And/or Menstrual Cycle PhasesJoaquim Guasch BoschNo ratings yet
- Menstrual Cycle Associated Changes in Agility in Amateur Female AthletesDocument39 pagesMenstrual Cycle Associated Changes in Agility in Amateur Female Athletesvishnu priyaNo ratings yet
- Translational Endocrinology of Bone: Reproduction, Metabolism, and the Central Nervous SystemFrom EverandTranslational Endocrinology of Bone: Reproduction, Metabolism, and the Central Nervous SystemGerard KarsentyNo ratings yet
- Ijerph 18 10024Document8 pagesIjerph 18 10024Tri AstutiNo ratings yet
- HHS Public Access: Estrogen-And Progesterone-Mediated Structural Neuroplasticity in Women: Evidence From NeuroimagingDocument40 pagesHHS Public Access: Estrogen-And Progesterone-Mediated Structural Neuroplasticity in Women: Evidence From NeuroimagingRoxana RoxNo ratings yet
- The Role of Physical Therapy in Sexual Health in Men and Women: Evaluation and TreatmentDocument11 pagesThe Role of Physical Therapy in Sexual Health in Men and Women: Evaluation and TreatmentMikeNo ratings yet
- Sleep and Neurologic DiseaseFrom EverandSleep and Neurologic DiseaseMitchell G. MiglisNo ratings yet
- A Case Study of Balance Rehabilitation in Parkinson's DiseaseDocument8 pagesA Case Study of Balance Rehabilitation in Parkinson's Diseaseddb5013No ratings yet
- The Role of Physical Therapy in Sexual Health in Men and WomenDocument11 pagesThe Role of Physical Therapy in Sexual Health in Men and WomenJorgeGalleguillosCavadaNo ratings yet
- In-Vivo Dynamics of The Human: Hippocampus Across The Menstrual CycleDocument9 pagesIn-Vivo Dynamics of The Human: Hippocampus Across The Menstrual CycleizaquenNo ratings yet
- The Influence of Yoga On Genetic Expression and EpigeneticsDocument3 pagesThe Influence of Yoga On Genetic Expression and Epigeneticscool MsA-LNo ratings yet
- Rehab September 2023Document8 pagesRehab September 2023Faeyz OrabiNo ratings yet
- Motivation To Eat and Not To Eat - The Psycho-Biological Conflict in Anorexia NervosaDocument6 pagesMotivation To Eat and Not To Eat - The Psycho-Biological Conflict in Anorexia NervosaCristina PaizNo ratings yet
- The Effect of Menstrual Cycle and Contraceptives On ACL Injuries and LaxityDocument10 pagesThe Effect of Menstrual Cycle and Contraceptives On ACL Injuries and Laxityxavi5_5No ratings yet
- Neri Journal R DutyDocument5 pagesNeri Journal R DutyAnamarcel Barillo NeriNo ratings yet
- Hormones and Embryonic Development: Advances in the BiosciencesFrom EverandHormones and Embryonic Development: Advances in the BiosciencesG. RaspéNo ratings yet
- Adrenal Disorders: Physiology, Pathophysiology and TreatmentFrom EverandAdrenal Disorders: Physiology, Pathophysiology and TreatmentAlice C. LevineNo ratings yet
- Accepted: Psychosomatic MedicineDocument24 pagesAccepted: Psychosomatic MedicineYekkit TinitanaNo ratings yet
- 2013 Shape Shifting PainDocument18 pages2013 Shape Shifting PainCristian OyarzoNo ratings yet
- Effects of Diaphragmatic Breathing Patterns On Balance: A Preliminary Clinical TrialDocument8 pagesEffects of Diaphragmatic Breathing Patterns On Balance: A Preliminary Clinical TrialDevasyaNo ratings yet
- Jung 2008Document5 pagesJung 2008pokharelriwaj82No ratings yet
- J American Geriatrics Society - 2016 - Spina - Music Therapy For Motor and Nonmotor Symptoms of Parkinson S Disease ADocument4 pagesJ American Geriatrics Society - 2016 - Spina - Music Therapy For Motor and Nonmotor Symptoms of Parkinson S Disease AFrancesco CiongoliNo ratings yet
- OA ESTROGEN Role of Sex Hormones in The Development of OsteoarthritisDocument5 pagesOA ESTROGEN Role of Sex Hormones in The Development of OsteoarthritisronniemonsterNo ratings yet
- Effect of Sex On MusclesDocument6 pagesEffect of Sex On Musclesapi-458763627No ratings yet
- Association Between Sleep Quality and Physical Activity in Postpartum Women - 2019Document8 pagesAssociation Between Sleep Quality and Physical Activity in Postpartum Women - 2019Franck BrardNo ratings yet
- 10 Su2016Document41 pages10 Su2016Sergio Machado NeurocientistaNo ratings yet
- The Effects of Exercise On Self-Rated Sleep Among Adults With Chronic Sleep ComplaintsDocument10 pagesThe Effects of Exercise On Self-Rated Sleep Among Adults With Chronic Sleep ComplaintsDanielaa ZapataNo ratings yet
- Greendale, 2009Document9 pagesGreendale, 2009Carolina EstradaNo ratings yet
- JCDR 6 1640Document4 pagesJCDR 6 1640saffrin syedNo ratings yet
- ShareefDocument12 pagesShareefParaschiva PîțuNo ratings yet
- The Effect of Pelvic Floor Muscle ExerciseDocument16 pagesThe Effect of Pelvic Floor Muscle Exercisesipen poltekkesbdgNo ratings yet
- Cholinergic Ligand InteractionsFrom EverandCholinergic Ligand InteractionsD. J. TriggleNo ratings yet
- Anatomy of ClitorisDocument2 pagesAnatomy of ClitorisOstrai OstraiNo ratings yet
- Research Article: International Journal of Recent Scientific Research Vol. 7, Issue, 11, Pp. 14342-14348, November, 2016Document7 pagesResearch Article: International Journal of Recent Scientific Research Vol. 7, Issue, 11, Pp. 14342-14348, November, 2016auliagyntrNo ratings yet
- Steroids Build More Muscles Without TrainingDocument10 pagesSteroids Build More Muscles Without TrainingLikith NagarajNo ratings yet
- 17141-Article Text-66770-1-10-20161211Document13 pages17141-Article Text-66770-1-10-20161211Hina IqbalNo ratings yet
- Berent-Spillson Et Al, 2015Document25 pagesBerent-Spillson Et Al, 2015Ezequiel NascimentoNo ratings yet
- Ejercicios Fernando Gomez PinillaDocument14 pagesEjercicios Fernando Gomez PinillaYajaira Saavedra JimenezNo ratings yet
- Academic Audience Final DraftDocument18 pagesAcademic Audience Final Draftapi-708250948No ratings yet
- Trends in Exercise Neuroscience: Raising Demand For Brain FitnessDocument4 pagesTrends in Exercise Neuroscience: Raising Demand For Brain FitnessfransAPNo ratings yet
- Papers StretchingDocument23 pagesPapers StretchingDiane Troncoso AlegriaNo ratings yet
- Randomized Controlled Trial of Yoga and Exercise in Multiple SclerosisDocument8 pagesRandomized Controlled Trial of Yoga and Exercise in Multiple Sclerosisfrogsmasher420No ratings yet
- Neurophysiological Effects of Spinal Manipulation: Joel G. Pickar, DC, PHDDocument15 pagesNeurophysiological Effects of Spinal Manipulation: Joel G. Pickar, DC, PHDAnonymous ZmRV6WqNo ratings yet
- Stress Inflammation & Yoga 2010Document9 pagesStress Inflammation & Yoga 2010Mike F MartelliNo ratings yet
- Bourke Harrell Neigh 2012 Stress GlucocorticoidsDocument20 pagesBourke Harrell Neigh 2012 Stress GlucocorticoidsConstanceNo ratings yet
- Nihms 123440Document19 pagesNihms 123440Gabriel CostaNo ratings yet
- Estrogen PsychikaDocument18 pagesEstrogen PsychikaKarina GardełoNo ratings yet
- Acupuncture IVFDocument10 pagesAcupuncture IVFWolfgang Rb100% (1)
- The Effect of Menstrual Cycle and Contraceptives On ACL Injuries and LaxityDocument10 pagesThe Effect of Menstrual Cycle and Contraceptives On ACL Injuries and Laxityxavi5_5No ratings yet
- Journal Pre-Proof: Journal of Pediatric and Adolescent GynecologyDocument3 pagesJournal Pre-Proof: Journal of Pediatric and Adolescent Gynecologyxavi5_5No ratings yet
- Walidd Score, A New Tool To Diagnose Dysmenorrhea and Predict Medical Leave in University StudentsDocument11 pagesWalidd Score, A New Tool To Diagnose Dysmenorrhea and Predict Medical Leave in University Studentsxavi5_5No ratings yet
- 17b-Estradiol Induced Effects On Anterior Cruciate Ligament Laxness and Neuromuscular Activation Patterns in Female RunnersDocument11 pages17b-Estradiol Induced Effects On Anterior Cruciate Ligament Laxness and Neuromuscular Activation Patterns in Female Runnersxavi5_5No ratings yet
- Balanchadar 2017Document11 pagesBalanchadar 2017xavi5_5No ratings yet
- Zazulak2006 PDFDocument16 pagesZazulak2006 PDFxavi5_5No ratings yet
- The Influence of Sex Hormones On Anterior Cruciate Ligament Rupture: Female StudyDocument8 pagesThe Influence of Sex Hormones On Anterior Cruciate Ligament Rupture: Female Studyxavi5_5No ratings yet
- The Menstrual Cycle May Affect Anterior Knee Laxity and The Rate of Anterior Cruciate Ligament RuptureDocument11 pagesThe Menstrual Cycle May Affect Anterior Knee Laxity and The Rate of Anterior Cruciate Ligament Rupturexavi5_5No ratings yet
- Seyed Esmaeel Shafiei2016Document4 pagesSeyed Esmaeel Shafiei2016xavi5_5No ratings yet
- Samuelson2017 PDFDocument13 pagesSamuelson2017 PDFxavi5_5No ratings yet
- LeBlanc 2017Document11 pagesLeBlanc 2017xavi5_5No ratings yet
- Textbook Proprioception in Orthopaedics Sports Medicine and Rehabilitation Defne Kaya Ebook All Chapter PDFDocument63 pagesTextbook Proprioception in Orthopaedics Sports Medicine and Rehabilitation Defne Kaya Ebook All Chapter PDFdaniel.ervin781100% (5)
- Reflex ResponseDocument10 pagesReflex Responseapi-264015665No ratings yet
- Reflexes ChapterDocument9 pagesReflexes ChapterVamsi KrishnaNo ratings yet
- Lecture 23 - Spinal Reflexes - KrakauerDocument2 pagesLecture 23 - Spinal Reflexes - Krakauersejal3vijNo ratings yet
- Muscle Tone PhysiologyDocument5 pagesMuscle Tone PhysiologyfatimaNo ratings yet
- Reflex ArcDocument8 pagesReflex ArcHina AsifNo ratings yet
- Structuring The Warm-Up: FASCIA AND SMR (Self-Myofascial Release)Document11 pagesStructuring The Warm-Up: FASCIA AND SMR (Self-Myofascial Release)ok okNo ratings yet
- Activitiy No. 12 - Human ReflexesDocument17 pagesActivitiy No. 12 - Human ReflexesTrix ThursdayNo ratings yet
- DocxDocument4 pagesDocxKimNo ratings yet
- ReflexesDocument35 pagesReflexesNadia AhmedNo ratings yet
- Golgi Tendon ReflexDocument15 pagesGolgi Tendon ReflexDemuel Dee L. BertoNo ratings yet
- Botulinum Toxin Therapeutic Mechanisms of Action.Document7 pagesBotulinum Toxin Therapeutic Mechanisms of Action.javierapobletevergaraNo ratings yet
- L1 Motor Function of CNSDocument48 pagesL1 Motor Function of CNSAhmed JawdetNo ratings yet
- Approach To Hypotonic InfantDocument56 pagesApproach To Hypotonic Infantar bindra100% (1)
- Krok1 PreparedDocument322 pagesKrok1 PreparedGMP EuropeNo ratings yet
- Neuroanatomy and Physiology of The Masticatory System: MAJ Joseph Lowe October 15, 2010Document76 pagesNeuroanatomy and Physiology of The Masticatory System: MAJ Joseph Lowe October 15, 2010joeylowetylerNo ratings yet
- Strain and Counterstrain For Structural IntegratorsDocument8 pagesStrain and Counterstrain For Structural IntegratorssimonyanNo ratings yet
- Reflexes: Involuntary Motor Response To A Sensory Stimulus Forms The Reflex ActionDocument33 pagesReflexes: Involuntary Motor Response To A Sensory Stimulus Forms The Reflex ActionbnmjgcNo ratings yet
- 07d - Spinal Cord - Reflexes - Ninja Nerd & DR NajeebDocument7 pages07d - Spinal Cord - Reflexes - Ninja Nerd & DR Najeebhiba jasimNo ratings yet
- Preeti Raghavan - Spasticity and Muscle Stiffness - Restoring Form and Function-Springer (2022)Document329 pagesPreeti Raghavan - Spasticity and Muscle Stiffness - Restoring Form and Function-Springer (2022)Renan Sequini FavaroNo ratings yet
- 1st MBBS PhysiologyDocument17 pages1st MBBS PhysiologyplsssssNo ratings yet
- Human Body Care Activity 5Document5 pagesHuman Body Care Activity 5Camila PrzNo ratings yet
- Reflex: DR Raghuveer ChoudharyDocument64 pagesReflex: DR Raghuveer ChoudharyPhysiology by Dr RaghuveerNo ratings yet
- © 2016 Pearson Education, IncDocument16 pages© 2016 Pearson Education, IncChristina GonezNo ratings yet
- Assessing The Reflexes Bell-Magendie LawDocument9 pagesAssessing The Reflexes Bell-Magendie LawRoger ViloNo ratings yet
- 3 Motor ExaminationDocument25 pages3 Motor ExaminationkikiNo ratings yet
- Stretch ReflexDocument31 pagesStretch ReflexTots Enriquez Navarro100% (1)
- REFLEXESDocument15 pagesREFLEXESSonali Soumyashree100% (2)
- A2 Buguina Peaches A.Document7 pagesA2 Buguina Peaches A.Peach Amparo BuguinaNo ratings yet
- Muscle Stretch ReflexDocument6 pagesMuscle Stretch ReflexNasreen SultanaNo ratings yet