You might also like
- Chapter Eleven - Pre Implantation Developmen - 2018 - Current Topics in DevelopmDocument28 pagesChapter Eleven - Pre Implantation Developmen - 2018 - Current Topics in DevelopmLudimilaNo ratings yet
- SPERMATOGENESIS Spermatids Zirkin and Goldberg 2018Document5 pagesSPERMATOGENESIS Spermatids Zirkin and Goldberg 2018LudimilaNo ratings yet
- SPERMATOGENESIS Spermatocytes Schimenti and Singh 2018Document6 pagesSPERMATOGENESIS Spermatocytes Schimenti and Singh 2018LudimilaNo ratings yet
- SPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018Document6 pagesSPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018LudimilaNo ratings yet
- Single Cell RNA Sequencing of The Cynomolgus Macaque Testis Re - 2020 - DevelopmDocument27 pagesSingle Cell RNA Sequencing of The Cynomolgus Macaque Testis Re - 2020 - DevelopmLudimilaNo ratings yet
- TESTIS Endocrinology Overview RSS Yuen Et Al 2018Document7 pagesTESTIS Endocrinology Overview RSS Yuen Et Al 2018LudimilaNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Jesus Hold My Hand EbDocument2 pagesJesus Hold My Hand EbGregg100% (3)
- Bio Lab Report GerminationDocument10 pagesBio Lab Report GerminationOli Damaskova100% (4)
- Horgolás Minta - PulcsiDocument5 pagesHorgolás Minta - PulcsiCagey Ice-RoyNo ratings yet
- Mythologia: PrologueDocument14 pagesMythologia: ProloguecentrifugalstoriesNo ratings yet
- Bike Chasis DesignDocument7 pagesBike Chasis Designparth sarthyNo ratings yet
- Saln 1994 FormDocument3 pagesSaln 1994 FormJulius RarioNo ratings yet
- Question 1 (1 Point) : SavedDocument31 pagesQuestion 1 (1 Point) : SavedCates TorresNo ratings yet
- Posthumanism Cyborgs and Interconnected Bodies by Jon BaileyDocument59 pagesPosthumanism Cyborgs and Interconnected Bodies by Jon BaileyDavid García MonteroNo ratings yet
- Problem Based LearningDocument23 pagesProblem Based Learningapi-645777752No ratings yet
- CO-PO MappingDocument6 pagesCO-PO MappingArun Kumar100% (1)
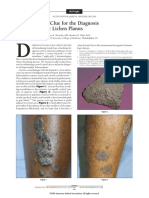
- A Novel Visual Clue For The Diagnosis of Hypertrophic Lichen PlanusDocument1 pageA Novel Visual Clue For The Diagnosis of Hypertrophic Lichen Planus600WPMPONo ratings yet
- Visual Acuity: Opthalmology CEX StepsDocument5 pagesVisual Acuity: Opthalmology CEX StepsVanessa HermioneNo ratings yet
- Succession CasesDocument17 pagesSuccession CasesAmbisyosa PormanesNo ratings yet
- Sauna Studies As An Academic Field: A New Agenda For International ResearchDocument42 pagesSauna Studies As An Academic Field: A New Agenda For International ResearchsedgehammerNo ratings yet
- Budget ProposalDocument1 pageBudget ProposalXean miNo ratings yet
- Agitha Diva Winampi - Childhood MemoriesDocument2 pagesAgitha Diva Winampi - Childhood MemoriesAgitha Diva WinampiNo ratings yet
- Task Performance Valeros Roeul GDocument6 pagesTask Performance Valeros Roeul GAnthony Gili100% (3)
- A Mercy Guided StudyDocument23 pagesA Mercy Guided StudyAnas HudsonNo ratings yet
- 2U6 S4HANA1909 Set-Up EN XXDocument10 pages2U6 S4HANA1909 Set-Up EN XXGerson Antonio MocelimNo ratings yet
- Danese and Romano (2011) ModerationDocument14 pagesDanese and Romano (2011) ModerationUmer NaseemNo ratings yet
- Law - Midterm ExamDocument2 pagesLaw - Midterm ExamJulian Mernando vlogsNo ratings yet
- Bakery Management SynopsisDocument13 pagesBakery Management SynopsisSHiVaM KRNo ratings yet
- AX Series Advanced Traffic Manager: Installation Guide For The AX 1030 / AX 3030Document18 pagesAX Series Advanced Traffic Manager: Installation Guide For The AX 1030 / AX 3030stephen virmwareNo ratings yet
- PH Water On Stability PesticidesDocument6 pagesPH Water On Stability PesticidesMontoya AlidNo ratings yet
- College PrepDocument2 pagesCollege Prepapi-322377992No ratings yet
- Judges Kings ProphetsDocument60 pagesJudges Kings ProphetsKim John BolardeNo ratings yet
- SBP Notes-1 PDFDocument7 pagesSBP Notes-1 PDFzeeshanNo ratings yet
- Civil Engineering Interview QuestionsDocument19 pagesCivil Engineering Interview QuestionsSrivardhanSrbNo ratings yet
- HUAWEI P8 Lite - Software Upgrade GuidelineDocument8 pagesHUAWEI P8 Lite - Software Upgrade GuidelineSedin HasanbasicNo ratings yet
- Pengaruh Abu Batu Sebagai Subtitusi Agregat Halus DanDocument10 pagesPengaruh Abu Batu Sebagai Subtitusi Agregat Halus Danangela merici rianawatiNo ratings yet