You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Effect of Oral Iron Administration On Mental State in Children With Low Serum Ferritin ConcentrationDocument5 pagesEffect of Oral Iron Administration On Mental State in Children With Low Serum Ferritin ConcentrationFrancisca AldunateNo ratings yet
- Perry 2006Document26 pagesPerry 2006Francisca AldunateNo ratings yet
- Jones2015 Translational Pyschiatry ConceptsDocument11 pagesJones2015 Translational Pyschiatry ConceptsFrancisca AldunateNo ratings yet
- Irnmunoneuroendocrine Role of Melatonin: Mini-ReviewDocument10 pagesIrnmunoneuroendocrine Role of Melatonin: Mini-ReviewFrancisca AldunateNo ratings yet
- Jun-13-SAHM VitaminD Position FinalDocument3 pagesJun-13-SAHM VitaminD Position FinalFrancisca AldunateNo ratings yet
- Issue Paper On COVID-19 and Fundamental Principles and Rights at WorkDocument38 pagesIssue Paper On COVID-19 and Fundamental Principles and Rights at WorkFrancisca AldunateNo ratings yet
- Melatonin As A Chronobiotic: Josephine Arendt, Debra Jean SkeneDocument15 pagesMelatonin As A Chronobiotic: Josephine Arendt, Debra Jean SkeneFrancisca AldunateNo ratings yet
- BCMJ 49 Vol3 Articles Posttraumatic StressDocument6 pagesBCMJ 49 Vol3 Articles Posttraumatic StressFrancisca AldunateNo ratings yet
- Hardeland 2005Document12 pagesHardeland 2005Francisca AldunateNo ratings yet
- Chanen Youth-SIG 15feb12Document40 pagesChanen Youth-SIG 15feb12Francisca AldunateNo ratings yet
- 7 Melatonin: Clinical Relevance: Russel J. ReiterDocument13 pages7 Melatonin: Clinical Relevance: Russel J. ReiterFrancisca AldunateNo ratings yet
- Alcohol Withdrawal Syndrome: Mechanisms, Manifestations, and ManagementDocument14 pagesAlcohol Withdrawal Syndrome: Mechanisms, Manifestations, and ManagementFrancisca AldunateNo ratings yet
- Childhood Traumas An Outline and Overview 1991 PDFDocument11 pagesChildhood Traumas An Outline and Overview 1991 PDFFrancisca AldunateNo ratings yet
- Treating OCD in Children and Adolescents: A Cognitive-Behavioral Approach Martin E. Franklin, PH.DDocument49 pagesTreating OCD in Children and Adolescents: A Cognitive-Behavioral Approach Martin E. Franklin, PH.DFrancisca AldunateNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Research Paper Topics On Child CareDocument7 pagesResearch Paper Topics On Child Careljkwfwgkf100% (1)
- Teenage PregnancyDocument3 pagesTeenage PregnancyCherrylou Buday100% (1)
- Developmental PsychologyDocument77 pagesDevelopmental PsychologyArkan KhairullahNo ratings yet
- MCN Lecture Midterms Reviewerpdf PDFDocument23 pagesMCN Lecture Midterms Reviewerpdf PDFRaphael Lorenz NavarroNo ratings yet
- Developmental PsychologyDocument3 pagesDevelopmental PsychologysdfghNo ratings yet
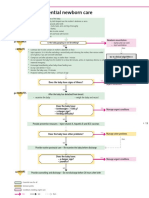
- Algorithm 2: Essential Newborn Care: BirthDocument1 pageAlgorithm 2: Essential Newborn Care: BirthGabriel Anthony ArtizaNo ratings yet
- Probable Defense QuestionsDocument2 pagesProbable Defense Questionscayla mae carlos100% (1)
- The Chinese Toddler Chained Through Love and FearDocument2 pagesThe Chinese Toddler Chained Through Love and FearTatiana KolesnikovaNo ratings yet
- Making Your Hospital Mother Baby FriendlyDocument19 pagesMaking Your Hospital Mother Baby Friendlydave100% (2)
- Chapter 2 of Teenage PregnancyDocument14 pagesChapter 2 of Teenage PregnancyHoney Joy Leopando SabadoNo ratings yet
- Ritual Life of The Igorots of Sagada, From Birth To AdolescenceDocument2 pagesRitual Life of The Igorots of Sagada, From Birth To AdolescenceEdison L. ChuNo ratings yet
- Pruhlen - What Was Best For An InfantDocument20 pagesPruhlen - What Was Best For An InfantSzekelyNo ratings yet
- This Study Resource Was: National University College Justicia CriminalDocument5 pagesThis Study Resource Was: National University College Justicia CriminalDavid RiveraNo ratings yet
- Being A Parent in The Real WorldDocument12 pagesBeing A Parent in The Real WorldBarbara BacauanuNo ratings yet
- RealCare Baby Op HandbookDocument51 pagesRealCare Baby Op HandbookFakeBabies100% (1)
- SP Final EssayDocument12 pagesSP Final Essayapi-660405105No ratings yet
- Read An Excerpt From Mayim Bialik's Beyond The Sling!Document18 pagesRead An Excerpt From Mayim Bialik's Beyond The Sling!Simon and Schuster100% (4)
- You Are NOT Alone!: Parents & IsolationDocument1 pageYou Are NOT Alone!: Parents & Isolationmickaylia greenNo ratings yet
- The Billionaire and The Nanny by North PaigeDocument291 pagesThe Billionaire and The Nanny by North PaigeSiphesihle NdlaziNo ratings yet
- Pre Discharge Scoring SheetDocument1 pagePre Discharge Scoring SheetdanielNo ratings yet
- Postnatal Care: Samson Udho MSC, BSCDocument34 pagesPostnatal Care: Samson Udho MSC, BSCAYO NELSON0% (1)
- The Magic War With Potch The MagicianDocument6 pagesThe Magic War With Potch The MagicianConsuelo Hurtado C.No ratings yet
- Kylah JacksonDocument5 pagesKylah Jacksonapi-548099066No ratings yet
- Baby PlannerDocument12 pagesBaby PlannerFattrin KartikasariNo ratings yet
- La Leche LeagueDocument5 pagesLa Leche Leagueapi-302732994No ratings yet
- Royal LoveDocument7 pagesRoyal LoveArianne DiosoNo ratings yet
- What To Do When The Boss Releases His Inner ToddlerDocument4 pagesWhat To Do When The Boss Releases His Inner ToddlerDUDUNG dudong100% (1)
- Child and Adolscnt 5Document18 pagesChild and Adolscnt 5Khiem RagoNo ratings yet
- (Ebook) Super Nanny SecretsDocument123 pages(Ebook) Super Nanny SecretsLoredana Cîrjan100% (1)
- Present Simple Versus Present ContinuousDocument15 pagesPresent Simple Versus Present ContinuousYogita BhamareNo ratings yet