You might also like
- Biopolym - Cell 2018 34 5 400 enDocument9 pagesBiopolym - Cell 2018 34 5 400 enАнна ШаповаловаNo ratings yet
- Flu UpdateDocument36 pagesFlu UpdateDan ShepardNo ratings yet
- Comparing Clinical Characteristics Between Hospitalized Adults With Laboratory-Confirmed Influenza A and B Virus InfectionDocument4 pagesComparing Clinical Characteristics Between Hospitalized Adults With Laboratory-Confirmed Influenza A and B Virus InfectionSofiaNo ratings yet
- PIIS0140673622009825Document14 pagesPIIS0140673622009825mauriciosagredoNo ratings yet
- Journal DermatologyDocument8 pagesJournal DermatologyImelva GirsangNo ratings yet
- Genomic Surveillance of COVID-19 Cases in Beijing: ArticleDocument9 pagesGenomic Surveillance of COVID-19 Cases in Beijing: ArticleCarlos CastroNo ratings yet
- h5n1 Research PapersDocument5 pagesh5n1 Research Papersafeaaawci100% (1)
- Dengue (Lancet)Document16 pagesDengue (Lancet)Roberth Mero100% (1)
- CDC Study On Effectiveness of COVID-19 VaccinesDocument8 pagesCDC Study On Effectiveness of COVID-19 VaccinesWSYX/WTTENo ratings yet
- J Infect Dis. 2016 Fonville 31 8Document8 pagesJ Infect Dis. 2016 Fonville 31 8Dwi Nur AfandiNo ratings yet
- Fimmu 05 00280Document10 pagesFimmu 05 00280Analia UrueñaNo ratings yet
- Tropicalmed 08 00016 v4Document11 pagesTropicalmed 08 00016 v4Shahazaman ShazuNo ratings yet
- A Multiplex Real-Time RT-PCR For Detection and Identification of Influenza Virus Types A and B AnDocument8 pagesA Multiplex Real-Time RT-PCR For Detection and Identification of Influenza Virus Types A and B AnMet RizalNo ratings yet
- PDF Jurding FikriDocument10 pagesPDF Jurding FikriAssifa RidzkiNo ratings yet
- Brote en Cordoba, Nuestro PaperDocument5 pagesBrote en Cordoba, Nuestro PaperClarabella FantilliNo ratings yet
- Volume 382, Issue Previous Article Next Article Cite or Link Using DOIDocument2 pagesVolume 382, Issue Previous Article Next Article Cite or Link Using DOIDAVIDSERGIO39946No ratings yet
- Pi Is 1473309921006484Document13 pagesPi Is 1473309921006484Leonardo IvrackNo ratings yet
- Dengue LancetDocument16 pagesDengue LancetJorge Ricardo Uchuya GómezNo ratings yet
- Estudio Publicado en Guangdong, China, Sobre La Contagiosidad de La Variante Delta de CoronavirusDocument18 pagesEstudio Publicado en Guangdong, China, Sobre La Contagiosidad de La Variante Delta de Coronavirussofia crottiNo ratings yet
- The Evolutionary Dynamics of Influenza A Virus Within and Between Human HostsDocument53 pagesThe Evolutionary Dynamics of Influenza A Virus Within and Between Human Hostsaleisha97No ratings yet
- Clinical Microbiology and Infection: Letter To The EditorDocument3 pagesClinical Microbiology and Infection: Letter To The EditorDiken LasimparaNo ratings yet
- HCV 3Document14 pagesHCV 3Mostafa ElashryNo ratings yet
- Influenza Vaccine Effectiveness and Statin Use Among Adults in The United States, 2011-2017Document7 pagesInfluenza Vaccine Effectiveness and Statin Use Among Adults in The United States, 2011-2017ullyfatmalaNo ratings yet
- JMV 24982Document9 pagesJMV 24982rks_rmrctNo ratings yet
- Nicholas J. Moss, Amalia Magaret, Kerry J. Laing, Angela Shaulov Kask, Minna Wang, Karen E. Mark, Joshua T. Schiffer, Anna Wald, and David M. KoelleDocument12 pagesNicholas J. Moss, Amalia Magaret, Kerry J. Laing, Angela Shaulov Kask, Minna Wang, Karen E. Mark, Joshua T. Schiffer, Anna Wald, and David M. KoelleIqaNo ratings yet
- Research Paper On h1n1 VirusDocument5 pagesResearch Paper On h1n1 Viruszijkchbkf100% (1)
- Influenza Activity - United States and Worldwide, 2007-08 SeasonDocument7 pagesInfluenza Activity - United States and Worldwide, 2007-08 SeasonjamesgarrowNo ratings yet
- Jiy103 - A Universal Influenza Vaccine The Strategic Plan For TheDocument8 pagesJiy103 - A Universal Influenza Vaccine The Strategic Plan For ThePGIndikaNo ratings yet
- Gilbert2005 Studi HivDocument12 pagesGilbert2005 Studi HivErnawati HidayatNo ratings yet
- Emerging Infectious Disease Agents and Their Potential Threat To Transfusion Safety.Document29 pagesEmerging Infectious Disease Agents and Their Potential Threat To Transfusion Safety.Pritha BhuwapaksophonNo ratings yet
- NIH Public Access: Author ManuscriptDocument12 pagesNIH Public Access: Author ManuscriptRiddhi GandhiNo ratings yet
- 8d9c PDFDocument11 pages8d9c PDFJulian AshbourneNo ratings yet
- Burnett2017Document16 pagesBurnett2017Jane DDNo ratings yet
- Coronavirus Inmunidad Cruzada Estudio de NigeriaDocument7 pagesCoronavirus Inmunidad Cruzada Estudio de NigeriaCarlos MendozaNo ratings yet
- Seroprevalence of Antibodies To Sars-Cov-2 in 10 Sites in The United States, March 23-May 12, 2020Document11 pagesSeroprevalence of Antibodies To Sars-Cov-2 in 10 Sites in The United States, March 23-May 12, 2020Mario Josue ValladaresGarridoNo ratings yet
- Mm7013e3 HDocument6 pagesMm7013e3 HRabin BaniyaNo ratings yet
- Increased Public Health Threat of Avian-Origin H3N2 Influenza Virus Caused by Its Evolution in DogsDocument21 pagesIncreased Public Health Threat of Avian-Origin H3N2 Influenza Virus Caused by Its Evolution in DogsNurul MaghfirohNo ratings yet
- WhoDocument7 pagesWhoYu YingNo ratings yet
- UkraineDocument6 pagesUkraineRasha MuhammadNo ratings yet
- Vassilopoulos 2002Document13 pagesVassilopoulos 2002deliaNo ratings yet
- 202 8 1154 PDFDocument7 pages202 8 1154 PDFSiti Zamilatul AzkiyahNo ratings yet
- Weekly Influenza Surveillance Report: During The Week Ending February 1, 2020Document6 pagesWeekly Influenza Surveillance Report: During The Week Ending February 1, 2020Timothy MooreNo ratings yet
- Bird Flu: MSC Epidemiology Doctor of Veterinary Medicine PhilippinesDocument33 pagesBird Flu: MSC Epidemiology Doctor of Veterinary Medicine PhilippinesYohanes FirmansyahNo ratings yet
- Transmission and Diagnosis of Human Immunodeficiency Virus Infection: An UpdateDocument2 pagesTransmission and Diagnosis of Human Immunodeficiency Virus Infection: An UpdateMiguel DiazNo ratings yet
- Oral Direct Acting Agent Therapy For Hepatitis C Virus Infection A Systematic ReviewDocument13 pagesOral Direct Acting Agent Therapy For Hepatitis C Virus Infection A Systematic ReviewRoberto López MataNo ratings yet
- Irv 12867Document10 pagesIrv 12867Ihsan fahrojiNo ratings yet
- Journal of Virology-2007-Szretter-2736.fullDocument9 pagesJournal of Virology-2007-Szretter-2736.fullilza millenia jaliusNo ratings yet
- El Riesgo de Las Infecciones Trasmitidas Por TransfusionDocument6 pagesEl Riesgo de Las Infecciones Trasmitidas Por TransfusionArturo sagasteguiNo ratings yet
- MMWR Weekly - July 23, 2021Document16 pagesMMWR Weekly - July 23, 2021Mark SchnyderNo ratings yet
- Swine Enteric Coronavirus Disease - A Review of 4 Years With Porcine Epidemic Diarrhoea Virus and Porcine Deltacoronavirus in The United States and CanadaDocument16 pagesSwine Enteric Coronavirus Disease - A Review of 4 Years With Porcine Epidemic Diarrhoea Virus and Porcine Deltacoronavirus in The United States and CanadaJuan Daniel Hernandez ValenzuelaNo ratings yet
- 2016 Article 1668Document7 pages2016 Article 1668Rahmiati LaoNo ratings yet
- Bacterial and Fungal Coinfections in Covid 19 Patients Hospitalized During The New York City Pandemic SurgeDocument5 pagesBacterial and Fungal Coinfections in Covid 19 Patients Hospitalized During The New York City Pandemic SurgeReynaldo CarvajalNo ratings yet
- Jix 244Document10 pagesJix 244Ricky SilaenNo ratings yet
- Mm7021e3 HDocument3 pagesMm7021e3 HShuvo H AhmedNo ratings yet
- Krammer2018 PDFDocument21 pagesKrammer2018 PDFJulio Martinez RodriguezNo ratings yet
- Dropulic 2019Document11 pagesDropulic 2019mostafaNo ratings yet
- Severe Acute Respiratory Syndrome Coronavirus 2 From Patient With Coronavirus Disease, United StatesDocument8 pagesSevere Acute Respiratory Syndrome Coronavirus 2 From Patient With Coronavirus Disease, United StatesLuis ricardo Véjar felixNo ratings yet
- Herpes Zoster Could Be An Early Manifestation of Undiagnosed Human Immunodeficiency Virus InfectionDocument5 pagesHerpes Zoster Could Be An Early Manifestation of Undiagnosed Human Immunodeficiency Virus InfectionMuhammad MaulanaNo ratings yet
- Articles: BackgroundDocument13 pagesArticles: BackgroundJoseph Adinolfi Jr.80% (5)
- Navigating The Microbial Basis of Inflammatory Bowel Diseases Seeing TheDocument7 pagesNavigating The Microbial Basis of Inflammatory Bowel Diseases Seeing TheFernando Ledesma BarrientosNo ratings yet
- J Infect Dis.-2016-Gaglani-1546-56 CompletoDocument25 pagesJ Infect Dis.-2016-Gaglani-1546-56 CompletoFernando Ledesma BarrientosNo ratings yet
- Anticuerpos Contra Dengue Aumentan ZikaDocument8 pagesAnticuerpos Contra Dengue Aumentan ZikaFernando Ledesma BarrientosNo ratings yet
- Anticuerpos Contra Dengue Aumentan ZikaDocument8 pagesAnticuerpos Contra Dengue Aumentan ZikaFernando Ledesma BarrientosNo ratings yet
- Anticuerpos Contra Dengue Aumentan ZikaDocument8 pagesAnticuerpos Contra Dengue Aumentan ZikaFernando Ledesma BarrientosNo ratings yet
- InTech Cultureconditionsandtypesofgrowthmediaformammaliancells PDFDocument16 pagesInTech Cultureconditionsandtypesofgrowthmediaformammaliancells PDFJonathan RedricoNo ratings yet
- Pediatric Vac + NotesDocument36 pagesPediatric Vac + NotesTrang VuNo ratings yet
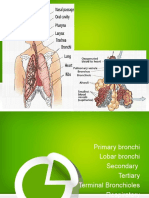
- Trans Pulmopatho ColoredDocument8 pagesTrans Pulmopatho Colored2012No ratings yet
- Nursing Care PlanDocument20 pagesNursing Care PlanmelliaiNo ratings yet
- Pedoman Nasional Pengendalian TuberkulosisDocument124 pagesPedoman Nasional Pengendalian Tuberkulosisykmadira serverNo ratings yet
- Nursing Care Plan For A Person With Croup Nursing DiagnosisDocument2 pagesNursing Care Plan For A Person With Croup Nursing DiagnosisMonica Rivera100% (1)
- NEJM 2014 Fundamentals of Lung AuscultationDocument7 pagesNEJM 2014 Fundamentals of Lung AuscultationFelipe CeaNo ratings yet
- 5 - Respiratory AssessmentDocument61 pages5 - Respiratory AssessmentAbboud Ali100% (1)
- 0 S02 - Ggn-Ii: Patientreportscsuperpanel - SP - General - Template01 - SC (Version: 7)Document2 pages0 S02 - Ggn-Ii: Patientreportscsuperpanel - SP - General - Template01 - SC (Version: 7)Yogesh BundelaNo ratings yet
- Parent Flu LetterDocument1 pageParent Flu LetterDennis BoxNo ratings yet
- RSKJikesiDocument9 pagesRSKJikesivonnyNo ratings yet
- PNEUMONIA and TubercolosisDocument5 pagesPNEUMONIA and TubercolosisAinee MeuvinNo ratings yet
- Sputum AnalysisDocument4 pagesSputum AnalysisGlenn PerezNo ratings yet
- Notes: Last Name First Name Middle Name Age Sex Address Contact NODocument108 pagesNotes: Last Name First Name Middle Name Age Sex Address Contact NOSample BakeshopNo ratings yet
- Health Declaration Form Health Declaration FormDocument1 pageHealth Declaration Form Health Declaration FormGerone NavaeraNo ratings yet
- Acute BronchitisDocument6 pagesAcute BronchitismelvinNo ratings yet
- GROUP 5 Some Facts and Theories About FluDocument14 pagesGROUP 5 Some Facts and Theories About FluRifat SebastianNo ratings yet
- NCP Ineffective Airway ClearanceDocument1 pageNCP Ineffective Airway ClearanceLucky RiusNo ratings yet
- DS SalbutamolDocument1 pageDS SalbutamolasdasdNo ratings yet
- Case Scenario - NebulizationDocument2 pagesCase Scenario - NebulizationHilario. Hayascent.Reign.M.No ratings yet
- Respiratory System QuestionsDocument8 pagesRespiratory System Questionschristine_8995No ratings yet
- Flu Symptoms: Flu Vaccine Each YearDocument2 pagesFlu Symptoms: Flu Vaccine Each YearKhristine ArellanoNo ratings yet
- Une Dyspnee Aigue Revelatrice Dun Syndrome de Mounier-Kuhn: A Propos Dun Cas Et Revue de LitteratureDocument6 pagesUne Dyspnee Aigue Revelatrice Dun Syndrome de Mounier-Kuhn: A Propos Dun Cas Et Revue de LitteratureIJAR JOURNALNo ratings yet
- Canine Influenza FactsDocument5 pagesCanine Influenza FactsWIS Digital News StaffNo ratings yet
- Lung Ultrasound Made Easy Step-By-Step GuideDocument40 pagesLung Ultrasound Made Easy Step-By-Step GuideJorge PérezNo ratings yet
- NIV Algorithm Final Version Jan 2020Document3 pagesNIV Algorithm Final Version Jan 2020AmalNo ratings yet
- Week 4 - Ms1 Course Task - Cu 4 - RoveroDocument2 pagesWeek 4 - Ms1 Course Task - Cu 4 - RoveroJohnmarkLorenzanaRoveroNo ratings yet
- BHP Formula No 08 Reference - Grindelia Pentarkan PTK 51 Drops For Bronchial Asthma - Homoeo - PK PDFDocument2 pagesBHP Formula No 08 Reference - Grindelia Pentarkan PTK 51 Drops For Bronchial Asthma - Homoeo - PK PDFDr. Kazy Habibur RahmanNo ratings yet
- Thorax and Lungs ReferenceDocument8 pagesThorax and Lungs Referencegwynth ripaldaNo ratings yet
- The Respiratory SystemDocument27 pagesThe Respiratory SystemmiryangelNo ratings yet
- Open Book Radiology Assesment: DR Zaleha Abd Manaf Hospital Pakar Al-IslamDocument28 pagesOpen Book Radiology Assesment: DR Zaleha Abd Manaf Hospital Pakar Al-IslamChua Kai XiangNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (24)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (80)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (1)
- Self-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!From EverandSelf-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!Rating: 5 out of 5 stars5/5 (1)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- To Explain the World: The Discovery of Modern ScienceFrom EverandTo Explain the World: The Discovery of Modern ScienceRating: 3.5 out of 5 stars3.5/5 (51)
- The Marshmallow Test: Mastering Self-ControlFrom EverandThe Marshmallow Test: Mastering Self-ControlRating: 4.5 out of 5 stars4.5/5 (58)