You might also like
- Practical 4: Cardiorespiratory Responses To Incremental ExerciseDocument4 pagesPractical 4: Cardiorespiratory Responses To Incremental ExercisePATRICK OTIATONo ratings yet
- Berbagai Macam Insulin Dan Cara KerjanyaDocument51 pagesBerbagai Macam Insulin Dan Cara KerjanyaAnonymous spBVX6t30UNo ratings yet
- Essential Oil For Blood Sugar BalanceDocument18 pagesEssential Oil For Blood Sugar BalanceNevaeh Valli Deeva100% (2)
- PT104 MidtermsDocument13 pagesPT104 MidtermsANGELI SIAOTONGNo ratings yet
- Changes in abdominal fat linked to weight loss and insulin sensitivityDocument7 pagesChanges in abdominal fat linked to weight loss and insulin sensitivityAres Santiago R. NoguedaNo ratings yet
- dirksen-et-al-2019-energy-intake-gastrointestinal-transit-and-gut-hormone-release-in-response-to-oral-triglycerides-and-2Document6 pagesdirksen-et-al-2019-energy-intake-gastrointestinal-transit-and-gut-hormone-release-in-response-to-oral-triglycerides-and-2Sulyaprilawati SiahaanNo ratings yet
- Intermittent Fasting and Refeeding On Insulin Dependant MenDocument10 pagesIntermittent Fasting and Refeeding On Insulin Dependant MenTyrone PenningNo ratings yet
- Physical Activity and Exercise in The Regulation of Human Adipose Tissue PhysiologyDocument35 pagesPhysical Activity and Exercise in The Regulation of Human Adipose Tissue PhysiologyLISTIA NURBAETINo ratings yet
- Aumento Plasmático Dos Poluentes Advém Da Lipólise Subcutânea - IJO - 01Document7 pagesAumento Plasmático Dos Poluentes Advém Da Lipólise Subcutânea - IJO - 01Elenice Peltz NutricionistaNo ratings yet
- Effect of Intermittent Fasting and Refeeding On Insulin Action in Healthy MenDocument9 pagesEffect of Intermittent Fasting and Refeeding On Insulin Action in Healthy Mendgh3No ratings yet
- Screenshot 2020-03-20 at 21.27.13Document3 pagesScreenshot 2020-03-20 at 21.27.13Denise KorkmazNo ratings yet
- Kiyoji Tanaka Maeng-Kyu Kim, Tsugio Tomita, Mi-Ji Kim, Hiroyuki Sasai, Seiji Maeda andDocument8 pagesKiyoji Tanaka Maeng-Kyu Kim, Tsugio Tomita, Mi-Ji Kim, Hiroyuki Sasai, Seiji Maeda andAnwar SiddiquiNo ratings yet
- Anabolizantes em Pacientes Pneumopatas 2Document7 pagesAnabolizantes em Pacientes Pneumopatas 2rayanna dornelesNo ratings yet
- Acute effects of microcurrent and exercise on abdominal fatDocument7 pagesAcute effects of microcurrent and exercise on abdominal fatNatalia Franco PérezNo ratings yet
- Is Visceral Adiposity A Significant Correlate of Subcutaneous Adipose Cell Lipolysis in Men?Document7 pagesIs Visceral Adiposity A Significant Correlate of Subcutaneous Adipose Cell Lipolysis in Men?Zachary LeeNo ratings yet
- tmpDDA TMPDocument7 pagestmpDDA TMPFrontiersNo ratings yet
- Spriet2014 Article NewInsightsIntoTheInteractionODocument10 pagesSpriet2014 Article NewInsightsIntoTheInteractionOaguaschma2114No ratings yet
- Effect of Low-And High-Glycemic-lndex Meals On Metabolism and Performance During High-Intensity, Intermittent ExerciseDocument11 pagesEffect of Low-And High-Glycemic-lndex Meals On Metabolism and Performance During High-Intensity, Intermittent ExerciseChris HaNo ratings yet
- Kanaley Aerobico SustratosDocument6 pagesKanaley Aerobico SustratosMartinVertolazNo ratings yet
- Fact #1: Wellness & Fitness-Body OptimizationDocument13 pagesFact #1: Wellness & Fitness-Body OptimizationCheeken CharliNo ratings yet
- Aquino Jr et al., 2012- versمo finalDocument10 pagesAquino Jr et al., 2012- versمo finalNoura RoseNo ratings yet
- The Glucose Fatty-Acid Cycle. Its Role In... (Randle1963) Randle CycleDocument5 pagesThe Glucose Fatty-Acid Cycle. Its Role In... (Randle1963) Randle CycleFilip ZaleskiNo ratings yet
- Effects of Dietary Fat On Muscle Substrates,.11Document9 pagesEffects of Dietary Fat On Muscle Substrates,.11Saul NavaNo ratings yet
- Tremblay-2011-Obesity The Allostatic Load of Weight Loss DietingDocument6 pagesTremblay-2011-Obesity The Allostatic Load of Weight Loss DietingluizfernandosellaNo ratings yet
- Dietary Protein Effect On Body CompositionDocument8 pagesDietary Protein Effect On Body CompositionAllisa RufaedahNo ratings yet
- New Perspectives On Nutritional Interventions To Augment Lipid Utilisation During ExerciseDocument12 pagesNew Perspectives On Nutritional Interventions To Augment Lipid Utilisation During ExerciseDani Prado GamboaNo ratings yet
- Art 3A10.1186 2Fs40795 017 0152 4Document9 pagesArt 3A10.1186 2Fs40795 017 0152 4NurfadhilahNo ratings yet
- 10 1152@japplphysiol 00907 2010Document11 pages10 1152@japplphysiol 00907 2010saulo freitas pereiraNo ratings yet
- Post Exercice Repletion of Muscle Energy Stores With Fructose or Glucose in Mixed MealsDocument9 pagesPost Exercice Repletion of Muscle Energy Stores With Fructose or Glucose in Mixed MealsLeonardo PinheiroNo ratings yet
- Thong Et Al. 2002 - Caffeine-Induced Impairment of Insulin Action But Not Insulin Signaling in HSM Is Reduced by ExerciseDocument8 pagesThong Et Al. 2002 - Caffeine-Induced Impairment of Insulin Action But Not Insulin Signaling in HSM Is Reduced by ExerciseAlbert CalvetNo ratings yet
- Obesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationDocument10 pagesObesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationRodrigoNo ratings yet
- Adiposa 2Document27 pagesAdiposa 2Nindya Agustin RNo ratings yet
- Cholesterol Metabolism and Body Composition in WomenDocument10 pagesCholesterol Metabolism and Body Composition in Womenpuspa raniNo ratings yet
- PIIS1550413118305655Document21 pagesPIIS1550413118305655Ilknur Okyay KellyNo ratings yet
- Aging Cell - 2021 - Chee - Increasing Skeletal Muscle Carnitine Content in Older Individuals Increases Whole Body FatDocument13 pagesAging Cell - 2021 - Chee - Increasing Skeletal Muscle Carnitine Content in Older Individuals Increases Whole Body FatBenjamínNo ratings yet
- Clinical Nutrition ESPEN: Original ArticleDocument8 pagesClinical Nutrition ESPEN: Original ArticleMar DianaNo ratings yet
- J. Lipid Res.-2007-Varady-2212-9Document8 pagesJ. Lipid Res.-2007-Varady-2212-9ggutierrezdieckNo ratings yet
- Philippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneDocument9 pagesPhilippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneMoonsferatuNo ratings yet
- The Influence of High-Carbohydrate Meals With Different Glycaemic Indices On Substrate UtilisationDocument8 pagesThe Influence of High-Carbohydrate Meals With Different Glycaemic Indices On Substrate UtilisationFer GominaNo ratings yet
- Eating Late Associated With Less Weight LossDocument8 pagesEating Late Associated With Less Weight Lossellla 38No ratings yet
- 2015 AjcnDocument8 pages2015 Ajcnseb2008No ratings yet
- Sedentarismo e Inflexibilidade MetabolicaDocument24 pagesSedentarismo e Inflexibilidade MetabolicaCristiano Kalata VarelaNo ratings yet
- Effect of Walking Exercise On Abdominal Fat, Insulin Resistance and Serum Cytokines in Obese WomenDocument9 pagesEffect of Walking Exercise On Abdominal Fat, Insulin Resistance and Serum Cytokines in Obese WomenankitsmkNo ratings yet
- Arad Et Al., 2015Document11 pagesArad Et Al., 2015Irene Chrysovalanto ThemistocleousNo ratings yet
- Cachexia-Inducing (MAC16) : Effect of Megestrol Weight by Factor and MiceDocument5 pagesCachexia-Inducing (MAC16) : Effect of Megestrol Weight by Factor and MicedonkeyendutNo ratings yet
- Fat and Carbohydrate Metabolism During Exercise in Elderly and Young SubjectsDocument7 pagesFat and Carbohydrate Metabolism During Exercise in Elderly and Young SubjectsMelati Noormaulidya PutriNo ratings yet
- Relationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidDocument10 pagesRelationship To Muscle Triglyceride and w-3 Fatty Acids in Muscle PhospholipidAdeess AengNo ratings yet
- Allman - 2019 OkDocument7 pagesAllman - 2019 OkMatheus SilvaNo ratings yet
- Exercise Training Induced Improved in Skeletal Muscle PGC-1Document9 pagesExercise Training Induced Improved in Skeletal Muscle PGC-1MTNC. César ZermeñoNo ratings yet
- Effects of Aerobic Exercise Performed in Fasted Versus Fed State On Fat and Carbohydrate..Document13 pagesEffects of Aerobic Exercise Performed in Fasted Versus Fed State On Fat and Carbohydrate..Salomé Horta LeytonNo ratings yet
- Articulo UciDocument11 pagesArticulo UciPEDRONo ratings yet
- Fasting REEDocument5 pagesFasting REEAimone FerriNo ratings yet
- Jejum IntermitenteDocument11 pagesJejum IntermitenteJohn Talmid De Yeshua HamashiachNo ratings yet
- ??? Metabolic SlowingDocument8 pages??? Metabolic SlowingJes BakerNo ratings yet
- Krista 2009Document9 pagesKrista 2009Vanessa BarrosNo ratings yet
- Ajcn 138214Document9 pagesAjcn 138214Gustavo AlmeidaNo ratings yet
- Moderate Weight Loss Attenuates Chronic Endoplasmic Reticulum Stress and Mitochondrial Dysfunction in Human ObesityDocument10 pagesModerate Weight Loss Attenuates Chronic Endoplasmic Reticulum Stress and Mitochondrial Dysfunction in Human ObesityJulia SCNo ratings yet
- Jurnal Lemak GoDocument9 pagesJurnal Lemak GoA09Aulia FebriantiNo ratings yet
- Cafeina y Ejercicio 2Document10 pagesCafeina y Ejercicio 2Rodrigo Durán San MartínNo ratings yet
- Fasting as Therapy - A Review of Intermittent Fasting Effects on HealthDocument12 pagesFasting as Therapy - A Review of Intermittent Fasting Effects on Healthlaiba ansariNo ratings yet
- Kiens 2011 Fat OxidationDocument7 pagesKiens 2011 Fat Oxidationlinoco1No ratings yet
- Jurnal FisikaDocument7 pagesJurnal FisikaKezia HanaraNo ratings yet
- Obesity - Short Scientific Findings to Ameliorate the Body WeightFrom EverandObesity - Short Scientific Findings to Ameliorate the Body WeightNo ratings yet
- Ni Hms 958771Document31 pagesNi Hms 958771Cem Tunaboylu (Student)No ratings yet
- Pros and Cons of Dietary Strategies Popular Among Cancer Patients - Cancer NetworkDocument1 pagePros and Cons of Dietary Strategies Popular Among Cancer Patients - Cancer NetworkCem Tunaboylu (Student)No ratings yet
- Pea Protein StudyDocument9 pagesPea Protein StudyCem Tunaboylu (Student)No ratings yet
- Pone 0209353 PDFDocument23 pagesPone 0209353 PDFFranklin Howley-Dumit SerulleNo ratings yet
- The Ketogenic Diet: Evidence For Optimism But High-Quality Research NeededDocument6 pagesThe Ketogenic Diet: Evidence For Optimism But High-Quality Research NeededCem Tunaboylu (Student)No ratings yet
- Rogers On 2017Document15 pagesRogers On 2017Paulo SCNo ratings yet
- Medicina: Nutritional Status and The Influence of The Vegan Diet On The Gut Microbiota and Human HealthDocument15 pagesMedicina: Nutritional Status and The Influence of The Vegan Diet On The Gut Microbiota and Human HealthCem Tunaboylu (Student)No ratings yet
- Flipping the metabolic switch: Understanding intermittent fastingDocument31 pagesFlipping the metabolic switch: Understanding intermittent fastingCem Tunaboylu (Student)No ratings yet
- Chester Et Al-2019-Diabetes Metabolism Research and ReviewsDocument10 pagesChester Et Al-2019-Diabetes Metabolism Research and ReviewsCem Tunaboylu (Student)No ratings yet
- Comparison-Based Computations Over Fully Homomorphic Encrypted DataDocument6 pagesComparison-Based Computations Over Fully Homomorphic Encrypted DataCem Tunaboylu (Student)No ratings yet
- Is Fasting Safe? A Chart Review of Adverse Events During Medically Supervised, Water-Only FastingDocument9 pagesIs Fasting Safe? A Chart Review of Adverse Events During Medically Supervised, Water-Only FastingRiley RilanNo ratings yet
- Alternate-Day Fasting in Nonobese Subjects: Effects On Body Weight, Body Composition, and Energy MetabolismDocument5 pagesAlternate-Day Fasting in Nonobese Subjects: Effects On Body Weight, Body Composition, and Energy MetabolismCem Tunaboylu (Student)No ratings yet
- Is Fasting Safe? A Chart Review of Adverse Events During Medically Supervised, Water-Only FastingDocument9 pagesIs Fasting Safe? A Chart Review of Adverse Events During Medically Supervised, Water-Only FastingRiley RilanNo ratings yet
- Adaptive Flight Control With Living Neuronal Networks On Microelectrode Trays - de Marse, Dockendorf PDFDocument4 pagesAdaptive Flight Control With Living Neuronal Networks On Microelectrode Trays - de Marse, Dockendorf PDFSubham SinghNo ratings yet
- Client-Side Encryption For Privacy-Sensitive Applications On The CloudDocument5 pagesClient-Side Encryption For Privacy-Sensitive Applications On The CloudCem Tunaboylu (Student)No ratings yet
- A Secure Database System Using Homomorphic Encryption SchemesDocument5 pagesA Secure Database System Using Homomorphic Encryption SchemesAsindu Wilfred DrilebaNo ratings yet
- Can Homomorphic Encryption Be Practical?Document18 pagesCan Homomorphic Encryption Be Practical?Cem Tunaboylu (Student)No ratings yet
- Building A Fully Homomorphic Encryption Scheme in PythonDocument14 pagesBuilding A Fully Homomorphic Encryption Scheme in PythonMuhammad AkramNo ratings yet
- Top foods herbs supplements reverse diabetes quicklyDocument7 pagesTop foods herbs supplements reverse diabetes quicklyMohammed ShazebNo ratings yet
- PDF 6Document8 pagesPDF 6Royyan AdiwijayaNo ratings yet
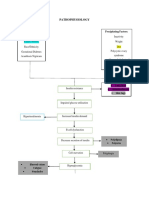
- Pathophysiology: Predisposing Factors: Precipitating FactorsDocument2 pagesPathophysiology: Predisposing Factors: Precipitating FactorsJemsMei Comparativo MensuradoNo ratings yet
- Academic Sciences International Journal of Pharmacy and Pharmaceutical SciencesDocument7 pagesAcademic Sciences International Journal of Pharmacy and Pharmaceutical SciencesEdy TansilNo ratings yet
- Benefits of Aerobic Exercise and RelatedDocument12 pagesBenefits of Aerobic Exercise and Relatedaleem_80No ratings yet
- Rohrbach, S 2007 AdiponeDocument35 pagesRohrbach, S 2007 AdiponeYHOISS SMIHT MUNOZ CERONNo ratings yet
- Milk Nutritional Composition and Its Role inDocument36 pagesMilk Nutritional Composition and Its Role inAzka FuadyNo ratings yet
- Everything You Need to Know About Intermittent Fasting (IFDocument8 pagesEverything You Need to Know About Intermittent Fasting (IFmay taurusNo ratings yet
- Da QingDocument18 pagesDa QingAlina PopaNo ratings yet
- DiabetesDocument15 pagesDiabetesSalman KhawarNo ratings yet
- Markers of B-Cell Failure Predict Poor Glycemic Response To GLP-1 Receptor Agonist Therapy in Type 2 DiabetesDocument8 pagesMarkers of B-Cell Failure Predict Poor Glycemic Response To GLP-1 Receptor Agonist Therapy in Type 2 DiabetesAnitaFauzziahYusufNo ratings yet
- Importance of Chromium for Dairy Cattle Milk ProductionDocument24 pagesImportance of Chromium for Dairy Cattle Milk ProductionGerardoNo ratings yet
- Diagnostic Criteria and Management of Metabolic Syndrome: Evolution OvertimeDocument26 pagesDiagnostic Criteria and Management of Metabolic Syndrome: Evolution OvertimeAjeng RetnoNo ratings yet
- Stop Sugar Addiction NOWDocument16 pagesStop Sugar Addiction NOWGgy VictorNo ratings yet
- Ensure Healthy Blood PressureDocument6 pagesEnsure Healthy Blood PressureWindy Ng WMNo ratings yet
- AAHA Diabetes Guidelines - FinalDocument21 pagesAAHA Diabetes Guidelines - FinalKátia SouzaNo ratings yet
- Pagt Tindakan BedahDocument33 pagesPagt Tindakan BedahRSUD PalmatakNo ratings yet
- Diabesity: A New Paradigm of Metabolic ConnexionDocument37 pagesDiabesity: A New Paradigm of Metabolic ConnexionatezorNo ratings yet
- Study Mango Leaves in Treating Type 2 DiabetesDocument10 pagesStudy Mango Leaves in Treating Type 2 DiabetesdragondostNo ratings yet
- Xylitol For HealthDocument24 pagesXylitol For HealthDonald WehmeyerNo ratings yet
- HypertriglyceridemiaDocument8 pagesHypertriglyceridemiaNaila KhanNo ratings yet
- Physiological Changes in PregnancyDocument40 pagesPhysiological Changes in PregnancyArhum SiddiqiNo ratings yet
- Review of Health Benefits of PeasDocument8 pagesReview of Health Benefits of PeasCrystal Joan D. MedranoNo ratings yet
- Diabetes Related DementiaDocument36 pagesDiabetes Related DementiaAndikhaNo ratings yet
- Metabolic response to injuryDocument5 pagesMetabolic response to injuryZllison Mae Teodoro Mangabat100% (1)
- DeFronzo PDFDocument23 pagesDeFronzo PDFMara HmNo ratings yet
- DIABETES ProjectDocument26 pagesDIABETES ProjectIshq KaurNo ratings yet