You might also like
- Neurology Multiple Choice Questions With Explanations: Volume IFrom EverandNeurology Multiple Choice Questions With Explanations: Volume IRating: 4 out of 5 stars4/5 (7)
- 0909 Case 2Document2 pages0909 Case 2willygopeNo ratings yet
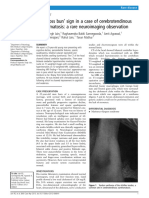
- Hot Cross Bun SignDocument4 pagesHot Cross Bun SignHenrique OliveiraNo ratings yet
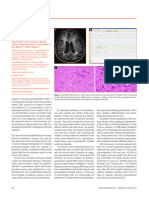
- Prion BrainDocument2 pagesPrion BrainD SNo ratings yet
- Apathy and Hypersomnia Are Common Features of Myotonic DystrophyDocument7 pagesApathy and Hypersomnia Are Common Features of Myotonic DystrophyAnnisa HusainNo ratings yet
- Hippocampal Granule Cell Necrosis in Untreated Addison's DiseaseDocument3 pagesHippocampal Granule Cell Necrosis in Untreated Addison's DiseaseJean Pierre Chastre LuzaNo ratings yet
- Proof of Progression Over Time Finally Fulminant Brain Muscle An 2009 SeiDocument3 pagesProof of Progression Over Time Finally Fulminant Brain Muscle An 2009 Seibilal hadiNo ratings yet
- Acute Kidney Injury and Proteinuria in A Man With HemoptysisDocument3 pagesAcute Kidney Injury and Proteinuria in A Man With HemoptysisChe GuevaraNo ratings yet
- 1175 FullDocument6 pages1175 FullAdnin NugrohoNo ratings yet
- BCR 2017 222185Document3 pagesBCR 2017 222185shofidhiaaaNo ratings yet
- Assessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationDocument8 pagesAssessment of The Distribution of Mitochondrial Ribosomal Rna in Melas and IN Thrombotic Cerebral Infarcts by in HybridizationVASCHOTI TRABANICONo ratings yet
- Del Toro 2006Document5 pagesDel Toro 2006NATALIA MARTINEZ CORDOBANo ratings yet
- Axonal Sensorimotor Neuropathy in Patients With B-ThalassemiaDocument5 pagesAxonal Sensorimotor Neuropathy in Patients With B-ThalassemiaDesty PurnamasariNo ratings yet
- Evidence of CNS Impairment in HIV Infection: Clinical, Neuropsychological, EEG, and MRI/MRS StudyDocument7 pagesEvidence of CNS Impairment in HIV Infection: Clinical, Neuropsychological, EEG, and MRI/MRS StudyLuther ThengNo ratings yet
- Electrophysiological Spectrum of Inclusion Body MyositisDocument3 pagesElectrophysiological Spectrum of Inclusion Body MyositisAmelito Miguel AndayaNo ratings yet
- Systemic Amyloidosis in Primary Hyperparathyroidism. Kam Newman, Mojtaba Akhtari, Salim Shackour, Ajit KesaniDocument1 pageSystemic Amyloidosis in Primary Hyperparathyroidism. Kam Newman, Mojtaba Akhtari, Salim Shackour, Ajit KesanijingerbrunoNo ratings yet
- Encefalitis LimbicaDocument11 pagesEncefalitis LimbicaRandy UlloaNo ratings yet
- Rare Case of Isolated Hypertrophic Neuropathy of the Oculomotor NerveDocument4 pagesRare Case of Isolated Hypertrophic Neuropathy of the Oculomotor NerveEvelin HarrizonNo ratings yet
- Multiple System Atrophy: Cellular and Molecular Pathology: D J Burn, E JarosDocument8 pagesMultiple System Atrophy: Cellular and Molecular Pathology: D J Burn, E Jarosuzair khanNo ratings yet
- BAB I OrangDocument7 pagesBAB I OrangDestriNo ratings yet
- NCP Neuro 0194Document5 pagesNCP Neuro 0194Fiaz medicoNo ratings yet
- Criteria For Diagnosis of Pure Neural Leprosy: Original CommunicationDocument4 pagesCriteria For Diagnosis of Pure Neural Leprosy: Original CommunicationFAZRI MONo ratings yet
- Cost of BC CareDocument7 pagesCost of BC CareErwin Chiquete, MD, PhDNo ratings yet
- Postictal Todd's Paralysis Associated with Focal Cerebral HypoperfusionDocument3 pagesPostictal Todd's Paralysis Associated with Focal Cerebral HypoperfusionNisa UcilNo ratings yet
- The New Phrenology The Limits of LocalizDocument2 pagesThe New Phrenology The Limits of LocalizgfrtbvhNo ratings yet
- Shimizu 2019Document7 pagesShimizu 2019jaimeaguilarr97No ratings yet
- Multiple-System AtrophyDocument15 pagesMultiple-System AtrophyNathasia SuryawijayaNo ratings yet
- Diagnosis, pathogenesis and treatment of primary episodic ataxiasDocument10 pagesDiagnosis, pathogenesis and treatment of primary episodic ataxiasthelegend 2022No ratings yet
- Incidental Complex Repetitive Discharges On Needle ElectromyographyDocument4 pagesIncidental Complex Repetitive Discharges On Needle Electromyographyfulgencio garciaNo ratings yet
- Adrenoleukodystrophy: Pathological StudyDocument15 pagesAdrenoleukodystrophy: Pathological StudyanniekrenNo ratings yet
- Adrenoleukodystrophy: Pathological StudyDocument15 pagesAdrenoleukodystrophy: Pathological StudyanniekrenNo ratings yet
- Acu 2017 1242Document14 pagesAcu 2017 1242Erin RyanNo ratings yet
- Ann MS Id 000849Document8 pagesAnn MS Id 000849azucenadeoNo ratings yet
- Magnetic Resonance Spectroscopy A Noninvasive Diagno - 2006 - Magnetic ResonancDocument3 pagesMagnetic Resonance Spectroscopy A Noninvasive Diagno - 2006 - Magnetic Resonanctejas1578No ratings yet
- What Are We Seeing?: Is Posterior Cortical Atrophy Just Alzheimer Disease?Document2 pagesWhat Are We Seeing?: Is Posterior Cortical Atrophy Just Alzheimer Disease?Eugenia FiginiNo ratings yet
- Isolated Propriospinal Myoclonus As A Presentation of Cervical Myelopathy.Document2 pagesIsolated Propriospinal Myoclonus As A Presentation of Cervical Myelopathy.arturomarticarvajalNo ratings yet
- Illustrative Teaching Case: Cerebral Fat EmbolismDocument3 pagesIllustrative Teaching Case: Cerebral Fat Embolismandy wookNo ratings yet
- Artigo Cientifico Charcot Marie ToothDocument8 pagesArtigo Cientifico Charcot Marie ToothCleverson Da Silva MattosNo ratings yet
- Chronic Immune Sensory Polyradiculopathy (CISP) : Stephanie Ong, Adam CassidyDocument4 pagesChronic Immune Sensory Polyradiculopathy (CISP) : Stephanie Ong, Adam Cassidylors93No ratings yet
- BH 010060 ENGDocument2 pagesBH 010060 ENGCarlosNo ratings yet
- Awards Works: Expanded AbstractDocument2 pagesAwards Works: Expanded AbstractJelvis BofNo ratings yet
- Hepaticka Encefalopatija I ManganDocument12 pagesHepaticka Encefalopatija I ManganIvan MihailovicNo ratings yet
- Posterior Cortical Dementia: Lost But Not ForgettingDocument2 pagesPosterior Cortical Dementia: Lost But Not ForgettingElisabet GobelliNo ratings yet
- Biologia Da Mielina e LeucodistrofiasDocument16 pagesBiologia Da Mielina e LeucodistrofiasFernando Cezar dos SantosNo ratings yet
- Neuroacanthocytosis SyndromesDocument9 pagesNeuroacanthocytosis SyndromesBreiner PeñarandaNo ratings yet
- Tuberculous Transverse Myelitis: Case Report and Review of The LiteratureDocument7 pagesTuberculous Transverse Myelitis: Case Report and Review of The LiteraturenoraNo ratings yet
- Effect Ofl Dopa On Oculogyric Crises inDocument2 pagesEffect Ofl Dopa On Oculogyric Crises inJoséNo ratings yet
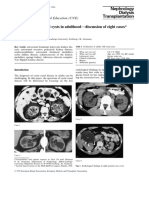
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- ATAXIAS PTDocument13 pagesATAXIAS PTKarunya VkNo ratings yet
- Algoritmo diferencial de la debilidad piramidal mas demenciaDocument6 pagesAlgoritmo diferencial de la debilidad piramidal mas demenciaFarid Santiago Abedrabbo LombeydaNo ratings yet
- Attempted Suicide by HangingDocument3 pagesAttempted Suicide by Hangingfarabi md.sharifuzzamanNo ratings yet
- Creutzfeldt JakobDocument15 pagesCreutzfeldt JakobNicolas RodriguezNo ratings yet
- A Case of Sporadic Creutzfeldt-JakobDocument5 pagesA Case of Sporadic Creutzfeldt-JakobVlad PopescuNo ratings yet
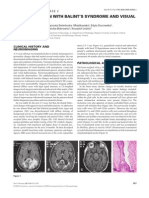
- 31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingDocument4 pages31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingwillygopeNo ratings yet
- Foster Kennedy Synd 2Document3 pagesFoster Kennedy Synd 2Anonymous k4F8sAs25No ratings yet
- Congenital Myasthenic Syndromes: Hometown HighlightsDocument3 pagesCongenital Myasthenic Syndromes: Hometown HighlightsApuse WakumNo ratings yet
- 全转录组分析肝性脑病小鼠海马区Document15 pages全转录组分析肝性脑病小鼠海马区GaryNo ratings yet
- Actinomycosis of Hip Complicated by Cns InfectionDocument4 pagesActinomycosis of Hip Complicated by Cns InfectionAmerNo ratings yet
- Rsob 180138Document19 pagesRsob 180138ilmiah neurologiNo ratings yet
- Strolling MatchDocument104 pagesStrolling MatchThienanNo ratings yet
- Altered Insula AnDocument9 pagesAltered Insula AnThienanNo ratings yet
- Gray Matter Decrease AnDocument8 pagesGray Matter Decrease AnThienanNo ratings yet
- Collecting and Applying Data On Social Determinants of Health in Health Care SettingsDocument4 pagesCollecting and Applying Data On Social Determinants of Health in Health Care SettingsThienanNo ratings yet
- Usher Syndrome CaseDocument4 pagesUsher Syndrome CaseThienanNo ratings yet
- Alt Brain Structure in AnDocument11 pagesAlt Brain Structure in AnThienanNo ratings yet
- On Eagle's Wings: MusicDocument16 pagesOn Eagle's Wings: MusicThienanNo ratings yet
- ECG Course Suggested Web SitesDocument1 pageECG Course Suggested Web SitesThienanNo ratings yet
- Neurobio of AnDocument15 pagesNeurobio of AnThienanNo ratings yet
- h4 Kf8 h5 Ke8 f7+ Kf8 Kf6 h6 Kg6 +-This Is The Key LineDocument1 pageh4 Kf8 h5 Ke8 f7+ Kf8 Kf6 h6 Kg6 +-This Is The Key LineThienanNo ratings yet
- Strolling MatchDocument104 pagesStrolling MatchThienanNo ratings yet
- Introduction To ECG ReadingDocument1 pageIntroduction To ECG ReadingThienanNo ratings yet
- Latex BibleDocument2 pagesLatex BibleLesPam Brenes100% (1)
- 2017 Toyota Highlander Product InfoDocument10 pages2017 Toyota Highlander Product InfoMateo DomínguezNo ratings yet
- Marshall AttackDocument1 pageMarshall AttackThienanNo ratings yet
- Schumann Ghost VariationsDocument9 pagesSchumann Ghost VariationsThienan100% (1)
- How To Practice Piano and Like It PDFDocument33 pagesHow To Practice Piano and Like It PDFfelipepaupitzNo ratings yet
- Janacek On An Overgrown PathDocument1 pageJanacek On An Overgrown PathThienanNo ratings yet
- Clarinet RepDocument13 pagesClarinet RepErick Álvarez-Sarabia80% (10)
- Haydn-Piano Sonate No.38Document10 pagesHaydn-Piano Sonate No.38Laci PatkoloNo ratings yet
- Manualonly 100602Document52 pagesManualonly 100602Orandi MoreiraNo ratings yet
- Schedule at The RestaurantDocument1 pageSchedule at The RestaurantThienanNo ratings yet
- Andrew Tang Interview: Insights from a Young Chess ProdigyDocument6 pagesAndrew Tang Interview: Insights from a Young Chess ProdigyThienanNo ratings yet
- Prayers of The FaithfulDocument1 pagePrayers of The FaithfulThienanNo ratings yet
- The Cold December El Desembre Congelat Piano SoloDocument3 pagesThe Cold December El Desembre Congelat Piano SoloAlexie Dagaerag MiraflorNo ratings yet
- Chess Interview: Thanks A Lot For Granting Me This Interview! Tell Us A Little About Yourself?Document8 pagesChess Interview: Thanks A Lot For Granting Me This Interview! Tell Us A Little About Yourself?ThienanNo ratings yet
- Effectiveness of Gallus Gallus Domesticus Eggshells As An Antibacterial Component of Hand SoapDocument71 pagesEffectiveness of Gallus Gallus Domesticus Eggshells As An Antibacterial Component of Hand SoapGabriel Christopher MembrilloNo ratings yet
- Deed of Undertaking Limited F2F InternshipDocument2 pagesDeed of Undertaking Limited F2F InternshipMarie LlanesNo ratings yet
- Nervous System ESSENTIAL QUESTIONSDocument8 pagesNervous System ESSENTIAL QUESTIONSAmanuelNo ratings yet
- Subcortical Ischemic Vascular Dementia: Helena C. Chui, MDDocument24 pagesSubcortical Ischemic Vascular Dementia: Helena C. Chui, MDEugenia FiginiNo ratings yet
- Thesis Final Last PDFDocument46 pagesThesis Final Last PDFDeepalNo ratings yet
- Health Education by Kamini MSNDocument15 pagesHealth Education by Kamini MSNkamini Choudhary100% (2)
- CNN International - Breaking News, US News, World News and VideoDocument8 pagesCNN International - Breaking News, US News, World News and VideomiculmucNo ratings yet
- Nursing responsibilities for montelukast sodiumDocument3 pagesNursing responsibilities for montelukast sodiumPrincess M Viznar BalolotNo ratings yet
- 00.docii 2020 Renal - UrogenitalDocument13 pages00.docii 2020 Renal - Urogenitalanderson ndabishakaNo ratings yet
- Schematic Pathophysiology CVADocument10 pagesSchematic Pathophysiology CVAheiyu100% (5)
- Acute Kidney Injury Concept MapDocument1 pageAcute Kidney Injury Concept MapKEn PilapilNo ratings yet
- Phlebotomy and Specimen ConsiderationsDocument21 pagesPhlebotomy and Specimen ConsiderationsLesly Marie LaxamanaNo ratings yet
- Applied Anatomy of Spinal CordDocument34 pagesApplied Anatomy of Spinal CordSuhana PathanNo ratings yet
- Literature Review FinalDocument4 pagesLiterature Review Finalapi-549244735No ratings yet
- WHO Insider Blows Whistle On Gates and GAVI - LewRockwellDocument9 pagesWHO Insider Blows Whistle On Gates and GAVI - LewRockwellZsi GaNo ratings yet
- Abdomen Macleod - S - Clinical - Examination - 15thDocument28 pagesAbdomen Macleod - S - Clinical - Examination - 15thwxyngtc4n9No ratings yet
- BIO PPT Presentation (1) 2Document28 pagesBIO PPT Presentation (1) 2luv.aryan879No ratings yet
- Tension Pneumothorax - StatPearls - NCBI BookshelfDocument9 pagesTension Pneumothorax - StatPearls - NCBI Bookshelfshinta damayantiNo ratings yet
- Relapse PreventionDocument38 pagesRelapse PreventionReshma Kiran100% (1)
- EY Employee Benefits ManualDocument49 pagesEY Employee Benefits ManualVishnu UdayagiriNo ratings yet
- Adverse Drug ReactionDocument7 pagesAdverse Drug ReactionufahadNo ratings yet
- 1.2 Listed DiseaseDocument23 pages1.2 Listed DiseaseplastiecoNo ratings yet
- Air Pollution Exposure and Cardiometabolic RiskDocument13 pagesAir Pollution Exposure and Cardiometabolic RiskWKYC.comNo ratings yet
- The Pathogenesis and Pathophysiology of Type 1 and Type 2 Diabetes MellitusDocument13 pagesThe Pathogenesis and Pathophysiology of Type 1 and Type 2 Diabetes MellitusKimberlyn CustodioNo ratings yet
- Drug Study - 5 DrugsDocument5 pagesDrug Study - 5 DrugsApril FlorendoNo ratings yet
- 05.012.002.0045-2 Finecare IiDocument2 pages05.012.002.0045-2 Finecare IiAinun Jariah100% (1)
- Proposed Regional Genitourinary Center: in CalabarzonDocument45 pagesProposed Regional Genitourinary Center: in Calabarzonallyssa monica duNo ratings yet
- Tim Dosen: Nur Rachmat, Bpo Cica Trimandasari N, Amd Op Muhammad Syaifuddin, Amd OpDocument11 pagesTim Dosen: Nur Rachmat, Bpo Cica Trimandasari N, Amd Op Muhammad Syaifuddin, Amd OpAdi NugrohoNo ratings yet
- Medical School of Ifakara (ST - Francs Refferal Hospital) : TH THDocument31 pagesMedical School of Ifakara (ST - Francs Refferal Hospital) : TH THMaryline WangweNo ratings yet
- Physical Education: DepedDocument67 pagesPhysical Education: DepedAj AntonioNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)