You might also like
- Central Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsFrom EverandCentral Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsNo ratings yet
- Cardio 2016Document14 pagesCardio 2016Mhmmd FasyaNo ratings yet
- Chapter 7 Regulation of Arterial Blood Pressure and MicrocirculationDocument20 pagesChapter 7 Regulation of Arterial Blood Pressure and Microcirculationaisyahasrii_No ratings yet
- Evaluation of Heart Rate Variability - UpToDateDocument31 pagesEvaluation of Heart Rate Variability - UpToDateGonçalo MirandaNo ratings yet
- How Baroreceptors Regulate Blood PressureDocument3 pagesHow Baroreceptors Regulate Blood PressureHaziq GanuNo ratings yet
- Chronic Interactions Between Carotid Baroreceptors and Chemoreceptors in Obesity HypertensionDocument14 pagesChronic Interactions Between Carotid Baroreceptors and Chemoreceptors in Obesity HypertensionCaesar Catalin CaratasuNo ratings yet
- Cardiovascular Changes in CardiogenicDocument4 pagesCardiovascular Changes in CardiogenicBaso AgusofyangNo ratings yet
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionRaam NaikNo ratings yet
- Artigo MargôDocument9 pagesArtigo MargôFredson André Nunes ValenteNo ratings yet
- Stochastic Model For Blood Pressure AnalysisDocument9 pagesStochastic Model For Blood Pressure AnalysisRakeshconclaveNo ratings yet
- Chronic Baroreflex Activation Restores Spontaneous Baroreflex Control and Variability of Heart Rate in Obesity-Induced HypertensionDocument10 pagesChronic Baroreflex Activation Restores Spontaneous Baroreflex Control and Variability of Heart Rate in Obesity-Induced HypertensionCaesar Catalin CaratasuNo ratings yet
- Acu CardioDocument13 pagesAcu CardioThais LimaNo ratings yet
- Heart Rate Variability PDFDocument7 pagesHeart Rate Variability PDFmik1989No ratings yet
- Cvs Group4Document14 pagesCvs Group4Lilian EdeniNo ratings yet
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionJoão Pedro Dantas AlkimimNo ratings yet
- Dehert 2012Document11 pagesDehert 2012putriNo ratings yet
- Cardiovascular Pathophysiology and Pharmacology NotesDocument6 pagesCardiovascular Pathophysiology and Pharmacology Notesjazzdoc007No ratings yet
- Revchilanestv50 04 16Document5 pagesRevchilanestv50 04 16ANA LAURA BRIONESNo ratings yet
- 1 Physiology and Pathophysiology of Thoracic Sympathetic BlockadeDocument7 pages1 Physiology and Pathophysiology of Thoracic Sympathetic BlockadeFarid AkbarNo ratings yet
- Nihms 1509292Document27 pagesNihms 1509292cloudnxneNo ratings yet
- The Human Sympathetic Nervous System: Its Relevance in Hypertension and Heart FailureDocument10 pagesThe Human Sympathetic Nervous System: Its Relevance in Hypertension and Heart FailureadeNo ratings yet
- Mechanisms Involved in Regulation of Systemic Blood PressureDocument6 pagesMechanisms Involved in Regulation of Systemic Blood PressureMumtaz MaulanaNo ratings yet
- Blood Pressure Regulation HandoutDocument10 pagesBlood Pressure Regulation Handoutsac50900100% (2)
- Autonomic DrugsDocument2 pagesAutonomic DrugsDouglas Greg CookNo ratings yet
- Meller2016 enDocument8 pagesMeller2016 encloudnxneNo ratings yet
- CVS QuesDocument8 pagesCVS QuesTan Ann JeeNo ratings yet
- Free-Diving - Control of Cerebral Blood FlowDocument5 pagesFree-Diving - Control of Cerebral Blood FlowTek StormanNo ratings yet
- Research Article: Yu Zheng, Jiping Zhang, Yanjie Wang, Yuying Wang, Yujun Lan, Shanshan Qu, Chunzhi Tang, and Yong HuangDocument10 pagesResearch Article: Yu Zheng, Jiping Zhang, Yanjie Wang, Yuying Wang, Yujun Lan, Shanshan Qu, Chunzhi Tang, and Yong Huangmazh21No ratings yet
- Cardiovascular Pathophysiology ExplainedDocument32 pagesCardiovascular Pathophysiology ExplainedSaif AliNo ratings yet
- The Rostral Ventral Lateral Medulla: Somato-Somatic ReflexesDocument2 pagesThe Rostral Ventral Lateral Medulla: Somato-Somatic ReflexesTrung Hiệp ĐàoNo ratings yet
- Cerebral Correlates of Autonomic Cardiovascular Arousal: A Functional Neuroimaging Investigation in HumansDocument12 pagesCerebral Correlates of Autonomic Cardiovascular Arousal: A Functional Neuroimaging Investigation in Humansbdalcin5512No ratings yet
- Hypertension: A Comparative Review Based On Fractal Wave Theory of ContinuumDocument8 pagesHypertension: A Comparative Review Based On Fractal Wave Theory of ContinuumAdaptive MedicineNo ratings yet
- HEARTRATEVARIABILITYDocument8 pagesHEARTRATEVARIABILITYdenta aeNo ratings yet
- 08 Autonomic Nervous SystemDocument0 pages08 Autonomic Nervous SystemjuniorebindaNo ratings yet
- Review Article: Regulation of Cerebral Blood FlowDocument8 pagesReview Article: Regulation of Cerebral Blood FlowAlmas TNo ratings yet
- Review Canine Idiopathic Dilated Cardiomyopathy. Part II: Pathophysiology and TherapyDocument14 pagesReview Canine Idiopathic Dilated Cardiomyopathy. Part II: Pathophysiology and TherapySarahNo ratings yet
- Controlled Hypotensive AnesthesiaDocument24 pagesControlled Hypotensive AnesthesiaSuresh Kumar100% (1)
- Anestesicos Locales y La Respuesta InflamatoriaDocument18 pagesAnestesicos Locales y La Respuesta InflamatoriarichabertNo ratings yet
- Blood Pressure RegulationDocument25 pagesBlood Pressure RegulationSyedzulqurnainhussainshah ZulqurnainNo ratings yet
- Blood Pressure Regulation SummaryDocument42 pagesBlood Pressure Regulation SummaryLouis JinNo ratings yet
- Cardiovascular and Autonomic Influences On Blood Pressure: John E. Jones,, Aruna R. Natarajan,,, and Pedro A. JoseDocument23 pagesCardiovascular and Autonomic Influences On Blood Pressure: John E. Jones,, Aruna R. Natarajan,,, and Pedro A. JoseLulu LuwiiNo ratings yet
- Autonomic Dysreflexia in Spinal Cord Injury: Causes, Symptoms, and TreatmentDocument14 pagesAutonomic Dysreflexia in Spinal Cord Injury: Causes, Symptoms, and Treatmentjacque zidaneNo ratings yet
- MJMHS 0626Document7 pagesMJMHS 0626lilydariniNo ratings yet
- Status Triphasicus Vs Status EpilepticusDocument8 pagesStatus Triphasicus Vs Status EpilepticusZaida RojasNo ratings yet
- Data Analysis and (1) (1)Document9 pagesData Analysis and (1) (1)khorshid aziziNo ratings yet
- Neuromodulation in Treatment of Hypertension by Acupuncture: A Neurophysiological ProspectiveDocument8 pagesNeuromodulation in Treatment of Hypertension by Acupuncture: A Neurophysiological Prospectivewillv4No ratings yet
- Blood Pressure Regulation - SlidesDocument35 pagesBlood Pressure Regulation - Slidesمحمد السنجلاويNo ratings yet
- Sample ChapterDocument14 pagesSample ChaptercpfredNo ratings yet
- Anatomy, Autonomic Nervous System - StatPearls - NCBI BookshelfDocument10 pagesAnatomy, Autonomic Nervous System - StatPearls - NCBI Bookshelfwood landerNo ratings yet
- Respiratory Physiology & NeurobiologyDocument6 pagesRespiratory Physiology & NeurobiologyCaroline BaschirottoNo ratings yet
- Lehrer2000 Article ResonantFrequencyBiofeedbackTrDocument15 pagesLehrer2000 Article ResonantFrequencyBiofeedbackTrAderilson OliveiraNo ratings yet
- Baroreceptor Modulation of The Cardiovascular System, Pain, Consciousness, and CognitionDocument51 pagesBaroreceptor Modulation of The Cardiovascular System, Pain, Consciousness, and CognitionPatoNo ratings yet
- Effects of periodic pulling on beating dynamics of cardiac tissueDocument20 pagesEffects of periodic pulling on beating dynamics of cardiac tissueSean ReabuyasNo ratings yet
- Sleep Disturbances in Highly Stress Reactive Mice: Modeling Endophenotypes of Major DepressionDocument13 pagesSleep Disturbances in Highly Stress Reactive Mice: Modeling Endophenotypes of Major DepressionYuriko AndreNo ratings yet
- Heart Rate Variability Analysis: Acta Physiologica Hungarica February 2001Document13 pagesHeart Rate Variability Analysis: Acta Physiologica Hungarica February 2001denta aeNo ratings yet
- Bondgraph Published CardioVascular 5Document24 pagesBondgraph Published CardioVascular 5AhmedNo ratings yet
- REGULATIONDocument19 pagesREGULATIONADITYAROOP PATHAKNo ratings yet
- Blood Pressure RegulationDocument35 pagesBlood Pressure Regulationبراءة أحمد السلاماتNo ratings yet
- Autonomic PharmacologyDocument14 pagesAutonomic Pharmacologyjanak019No ratings yet
- Reserch VestDocument11 pagesReserch VestRAMO STEF SZEKERESNo ratings yet
- Multicentre Analysis CancerDocument8 pagesMulticentre Analysis Cancersilvio da costa guerreiroNo ratings yet
- Resistance Exercise Dosage in Men With Prostate CancerDocument60 pagesResistance Exercise Dosage in Men With Prostate Cancersilvio da costa guerreiroNo ratings yet
- The Optimal Diet For Women With Polycystic Ovary SyndromeDocument12 pagesThe Optimal Diet For Women With Polycystic Ovary Syndromesilvio da costa guerreiroNo ratings yet
- Weight - Loss For Obese - Prostate - Cancer - Patients OnDocument9 pagesWeight - Loss For Obese - Prostate - Cancer - Patients Onsilvio da costa guerreiroNo ratings yet
- Nutritional Protocol With OncologyDocument14 pagesNutritional Protocol With Oncologysilvio da costa guerreiroNo ratings yet
- Nutrition Advanced Cancer CachexiaDocument9 pagesNutrition Advanced Cancer Cachexiasilvio da costa guerreiroNo ratings yet
- Exercise - Intolerance in Cystic FibrosisDocument10 pagesExercise - Intolerance in Cystic Fibrosissilvio da costa guerreiroNo ratings yet
- Exercise - Mode - Specificity - For - Preserving - Spine - and Cancer ProstateDocument8 pagesExercise - Mode - Specificity - For - Preserving - Spine - and Cancer Prostatesilvio da costa guerreiroNo ratings yet
- Protein - Supplementation - and - Resistance - Training in CancerDocument9 pagesProtein - Supplementation - and - Resistance - Training in Cancersilvio da costa guerreiroNo ratings yet
- Inflammation - Mediates - Exercise - Effects - On - Fatigue CancerDocument45 pagesInflammation - Mediates - Exercise - Effects - On - Fatigue Cancersilvio da costa guerreiroNo ratings yet
- Industry corner: rapid development of CDK4/6 inhibitor ribociclib for advanced breast cancerDocument4 pagesIndustry corner: rapid development of CDK4/6 inhibitor ribociclib for advanced breast cancersilvio da costa guerreiroNo ratings yet
- Low Skeletal Muscle Predictor Complications CancerDocument14 pagesLow Skeletal Muscle Predictor Complications Cancersilvio da costa guerreiroNo ratings yet
- Aerobic Recovery After Radical Prostatectomy: A Case StudyDocument7 pagesAerobic Recovery After Radical Prostatectomy: A Case Studysilvio da costa guerreiroNo ratings yet
- Jurnal 5Document9 pagesJurnal 5raraNo ratings yet
- American College of Gastroenterology Guideline .6Document16 pagesAmerican College of Gastroenterology Guideline .6Alex Antony Fernandez QuisocalaNo ratings yet
- Carotene in The Human Body MetabolicDocument20 pagesCarotene in The Human Body Metabolicsilvio da costa guerreiroNo ratings yet
- Sedentary - Behavior, Cadence and Physical Activity KneeDocument9 pagesSedentary - Behavior, Cadence and Physical Activity Kneesilvio da costa guerreiroNo ratings yet
- Clinical and Predictive Cancer PatientDocument13 pagesClinical and Predictive Cancer Patientsilvio da costa guerreiroNo ratings yet
- Traditional and New - Perspectives On Youth CardioDocument11 pagesTraditional and New - Perspectives On Youth Cardiosilvio da costa guerreiroNo ratings yet
- Thermoregulatory - Responses - To - Constant Versus Variables Intensity ExerciseDocument8 pagesThermoregulatory - Responses - To - Constant Versus Variables Intensity Exercisesilvio da costa guerreiroNo ratings yet
- A T2T Update in Ulcerative Colitis PDFDocument10 pagesA T2T Update in Ulcerative Colitis PDFmelvinNo ratings yet
- E B F R: Xercise and Lood LOW EstrictionDocument13 pagesE B F R: Xercise and Lood LOW Estrictionsilvio da costa guerreiroNo ratings yet
- The - Mechanoreflex and Hemodynamic - Response Passive LegDocument9 pagesThe - Mechanoreflex and Hemodynamic - Response Passive Legsilvio da costa guerreiroNo ratings yet
- Repeated - Sprints - Alter - Signaling - Related To MitochondrialDocument8 pagesRepeated - Sprints - Alter - Signaling - Related To Mitochondrialsilvio da costa guerreiroNo ratings yet
- Effect of HIIT On Remodeling Reumatoid ArthritisDocument9 pagesEffect of HIIT On Remodeling Reumatoid Arthritissilvio da costa guerreiroNo ratings yet
- The - Indirect Role of Executive - Functions On The CardiorrespiratotioDocument40 pagesThe - Indirect Role of Executive - Functions On The Cardiorrespiratotiosilvio da costa guerreiroNo ratings yet
- The - Role of Aerobic - Exercise in ReducingDocument6 pagesThe - Role of Aerobic - Exercise in Reducingsilvio da costa guerreiroNo ratings yet
- Estimation Model Cardiorespiratory Fitness ArthritisDocument44 pagesEstimation Model Cardiorespiratory Fitness Arthritissilvio da costa guerreiroNo ratings yet
- Energy Flow Analysis To Investigate Youth PitchingDocument9 pagesEnergy Flow Analysis To Investigate Youth Pitchingsilvio da costa guerreiroNo ratings yet
- Dynamic - Cerebral - Autoregulation Is MaintainedDocument7 pagesDynamic - Cerebral - Autoregulation Is Maintainedsilvio da costa guerreiroNo ratings yet
- Human Physiology From Cells To Systems 7Th Edition Sherwood Test Bank Cells To Systems 7Th Edition Sherwood Test Bank Full Chapter PDFDocument68 pagesHuman Physiology From Cells To Systems 7Th Edition Sherwood Test Bank Cells To Systems 7Th Edition Sherwood Test Bank Full Chapter PDFcleopatrabanhft1vh7100% (9)
- PHS 205 Cardiovascular SystemDocument33 pagesPHS 205 Cardiovascular Systemdivineraymond34No ratings yet
- Chapter 21 of Anatomy and Physiology by John Wiley and SonsDocument48 pagesChapter 21 of Anatomy and Physiology by John Wiley and SonsRonaldo ZapateroNo ratings yet
- Regulate Cardiac Output FactorsDocument49 pagesRegulate Cardiac Output FactorsPeter Mukunza100% (1)
- Reflexology AssignmentDocument5 pagesReflexology AssignmentelemibodyworksNo ratings yet
- Process of urine formation and blood pressure regulationDocument3 pagesProcess of urine formation and blood pressure regulationSamia TazinNo ratings yet
- #05 Autonomic Pharmacology v2Document122 pages#05 Autonomic Pharmacology v2tasnemaldaherNo ratings yet
- 15 PharmacologyDocument15 pages15 PharmacologySpy HanNo ratings yet
- Blood Pressure RegulationDocument11 pagesBlood Pressure RegulationPaulShaneHerreraZorrillaNo ratings yet
- Regulation of Heart RateDocument30 pagesRegulation of Heart RateTripat Singh67% (3)
- Pcol Cover To CoverDocument214 pagesPcol Cover To CoverJec OcampoNo ratings yet
- BZJ Reflex ReviewDocument11 pagesBZJ Reflex Reviewnarasaiyanhariharan2No ratings yet
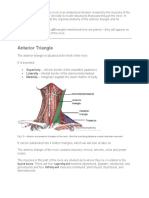
- The Anterior and Posterior Triangles of The NeckDocument10 pagesThe Anterior and Posterior Triangles of The NeckClair TomNo ratings yet
- Introduction To Physiology Eden - Sikoma XXXXDocument51 pagesIntroduction To Physiology Eden - Sikoma XXXXMarvelous MainzaNo ratings yet
- Unit 06 Blood VesselsDocument101 pagesUnit 06 Blood VesselsJohnykutty JosephNo ratings yet
- 18 Vessels and Flow DynamicsDocument57 pages18 Vessels and Flow Dynamicsraanja2No ratings yet
- Drugs Acting in CVSDocument63 pagesDrugs Acting in CVSMeghan Norico Cristuta100% (1)
- SAQ - OxytocinDocument3 pagesSAQ - Oxytocinanaeshkl100% (1)
- Cardiac Output, Blood Flow, and Blood PressureDocument69 pagesCardiac Output, Blood Flow, and Blood PressureUzma KhanNo ratings yet
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionRaam NaikNo ratings yet
- Cardiac ReflexesDocument2 pagesCardiac ReflexesfeliciaNo ratings yet
- Heart QuizDocument1 pageHeart QuizNaNo ratings yet
- Valsalva ManeuverDocument9 pagesValsalva ManeuverRina YulianaNo ratings yet
- Lecture Powerpoint Major Themes of Anatomy and PhysiologyDocument146 pagesLecture Powerpoint Major Themes of Anatomy and PhysiologykebNo ratings yet
- Circulatory System Guide: Blood Vessels, Circulation PhysiologyDocument45 pagesCirculatory System Guide: Blood Vessels, Circulation PhysiologyLloyd RebusiNo ratings yet
- JV Heart Rate, Heart Sounds PDFDocument39 pagesJV Heart Rate, Heart Sounds PDFRiya GuptaNo ratings yet
- Extrinsic Regulation of The HeartDocument9 pagesExtrinsic Regulation of The Heartkawaiijanai gfNo ratings yet
- My Lecture13 - Cardiovascular PhysiologyDocument16 pagesMy Lecture13 - Cardiovascular PhysiologyVivek ChaudharyNo ratings yet
- Blood Vessels CH 13Document86 pagesBlood Vessels CH 13Nalla Mirelle CarbonellNo ratings yet
- Homeostasis of The Cardiovascular SystemDocument7 pagesHomeostasis of The Cardiovascular SystemdesbestNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (14)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisFrom EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisRating: 5 out of 5 stars5/5 (8)