You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Nature326-0529-2016 Estres Reticulo EndoplasmicoDocument10 pagesNature326-0529-2016 Estres Reticulo EndoplasmicoErika Elena Zagal HuertaNo ratings yet
- RajamohanDocument7 pagesRajamohanErika Elena Zagal HuertaNo ratings yet
- RajamohanDocument7 pagesRajamohanErika Elena Zagal HuertaNo ratings yet
- Cinet Enz Wood - Metodos GraficosDocument6 pagesCinet Enz Wood - Metodos GraficosErika Elena Zagal HuertaNo ratings yet
- Romanowsky Citol Claus 11Document12 pagesRomanowsky Citol Claus 11Erika Elena Zagal HuertaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Laboratory Report No.1Document6 pagesLaboratory Report No.1mary hemarNo ratings yet
- Trump ReportDocument237 pagesTrump ReportAaron Nobel100% (1)
- O/W: Mayne To Reward The Willing: Mayne Pharma Group (MYX)Document8 pagesO/W: Mayne To Reward The Willing: Mayne Pharma Group (MYX)Muhammad ImranNo ratings yet
- "Mode One" Author & Dating Coach Alan Roger Currie Releases Criticism of Alleged Harvey Weinstein BehaviorDocument3 pages"Mode One" Author & Dating Coach Alan Roger Currie Releases Criticism of Alleged Harvey Weinstein BehaviorPR.com100% (1)
- DMD 2018 Stem and Non Stem Checklist Effective 2020-2021Document8 pagesDMD 2018 Stem and Non Stem Checklist Effective 2020-2021Elle DyNo ratings yet
- MahuaDocument12 pagesMahuaVinay ChhalotreNo ratings yet
- Early Theories On The Origin of Life On Earth: Creation TheoryDocument1 pageEarly Theories On The Origin of Life On Earth: Creation TheoryAngel Elizaga IINo ratings yet
- Drew Marine Chemical ManualDocument36 pagesDrew Marine Chemical ManualcpiquiNo ratings yet
- Two-Post Lift Model:210C 210CXDocument36 pagesTwo-Post Lift Model:210C 210CXventitapostNo ratings yet
- Recipes For Ceviche, Pebre and Pisco SourDocument4 pagesRecipes For Ceviche, Pebre and Pisco SourJoaquín Rolón CarrerasNo ratings yet
- Tinda Tomato PachadiDocument1 pageTinda Tomato PachadijyosriNo ratings yet
- Benign Prostate Hyperplasia 2Document125 pagesBenign Prostate Hyperplasia 2Danieal NeymarNo ratings yet
- Tolerance Chart: (Maximum Permissible Error)Document3 pagesTolerance Chart: (Maximum Permissible Error)arif_setyaw4nNo ratings yet
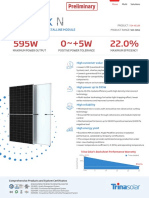
- DT-M Datasheet Vertex Ne19r en 2022 Pa1 Web Ne19r 2022pa1 en 20220916Document2 pagesDT-M Datasheet Vertex Ne19r en 2022 Pa1 Web Ne19r 2022pa1 en 20220916Producto ITICCOLNo ratings yet
- Daftar Pustaka ProposalDocument4 pagesDaftar Pustaka ProposalraniNo ratings yet
- Construction Quality Control PlanDocument35 pagesConstruction Quality Control PlanMalik Riaz100% (4)
- Farmall 105u115u 16x16transDocument2,233 pagesFarmall 105u115u 16x16transchrideerNo ratings yet
- Physical, Chemical and Biological HazardsDocument32 pagesPhysical, Chemical and Biological HazardsDaisy ObisoNo ratings yet
- Selection and Ranking of Rail Vehicle Components For Optimal Lightweighting Using Composite MaterialsDocument14 pagesSelection and Ranking of Rail Vehicle Components For Optimal Lightweighting Using Composite MaterialsAwan AJaNo ratings yet
- Material Safety Data Sheet Glyphosate 5.4Document5 pagesMaterial Safety Data Sheet Glyphosate 5.4Ahfi Rizqi FajrinNo ratings yet
- Cell Reproduction Practice ExamDocument5 pagesCell Reproduction Practice Examjacky qianNo ratings yet
- Calcium Cyanide: Hydrogen Cyanide. CONSULT THE NEW JERSEYDocument6 pagesCalcium Cyanide: Hydrogen Cyanide. CONSULT THE NEW JERSEYbacabacabacaNo ratings yet
- Chapter 4Document6 pagesChapter 4Aung MyatNo ratings yet
- Carbonyl Compounds: Unit 4 Rates, Equilibria and Further Organic ChemistryDocument5 pagesCarbonyl Compounds: Unit 4 Rates, Equilibria and Further Organic ChemistryKirusaanth SathiyathasanNo ratings yet
- Epoxy Plus Pipeline Repai Grout TG - MSDSDocument5 pagesEpoxy Plus Pipeline Repai Grout TG - MSDSgrimaguilNo ratings yet
- Manoshe Street Takeaway MenuDocument9 pagesManoshe Street Takeaway MenuimaddakrNo ratings yet
- Fuel EnergizerDocument17 pagesFuel Energizerrakshak25100% (1)
- TZ150-03 Service Manual - ChopperDocument42 pagesTZ150-03 Service Manual - ChopperMax Dean Araújo FernandesNo ratings yet
- Method Statement For Interlocking PavingDocument5 pagesMethod Statement For Interlocking Pavingdinidu3priyankara3saNo ratings yet
- Cocoa Nursery Manual Feb 2013Document51 pagesCocoa Nursery Manual Feb 2013jeffreyNo ratings yet