You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- 1993 - Linolenic Acid Flaxseed (Linum Usitatissimum)Document12 pages1993 - Linolenic Acid Flaxseed (Linum Usitatissimum)Geanelly SALGADO URIOSTEGUINo ratings yet
- 2020 - Draft Genome of Meyerozyma GuilliermondiiDocument6 pages2020 - Draft Genome of Meyerozyma GuilliermondiiGeanelly SALGADO URIOSTEGUINo ratings yet
- 2020 - Cyanobacterial PolyhydroxyalkanoatesDocument24 pages2020 - Cyanobacterial PolyhydroxyalkanoatesGeanelly SALGADO URIOSTEGUINo ratings yet
- YSI 5300A Biological Oxygen Monitor Manual ADocument48 pagesYSI 5300A Biological Oxygen Monitor Manual AGeanelly SALGADO URIOSTEGUINo ratings yet
- 1981 - Immunological Comparison of Ribosomal Proteins FromDocument7 pages1981 - Immunological Comparison of Ribosomal Proteins FromGeanelly SALGADO URIOSTEGUINo ratings yet
- 2020 - Outdoor Cultivation of Chlorella Sorokiniana in Third GenerationDocument7 pages2020 - Outdoor Cultivation of Chlorella Sorokiniana in Third GenerationGeanelly SALGADO URIOSTEGUINo ratings yet
- 2020 - Effect of Iron Oxide Nanoparticles On Growth and Biofuel Potential ofDocument11 pages2020 - Effect of Iron Oxide Nanoparticles On Growth and Biofuel Potential ofGeanelly SALGADO URIOSTEGUINo ratings yet
- Krebs Cycle Anions in Metabolic Acidosis: CommentaryDocument2 pagesKrebs Cycle Anions in Metabolic Acidosis: CommentaryGeanelly SALGADO URIOSTEGUINo ratings yet
- 1991 - Metabolism of (2-14C) Acetate and Its Use in Assessing Hepatic KrebDocument6 pages1991 - Metabolism of (2-14C) Acetate and Its Use in Assessing Hepatic KrebGeanelly SALGADO URIOSTEGUINo ratings yet
- 01 2006 - Succinate Dehydrogenase and Fumarate HydrataseDocument8 pages01 2006 - Succinate Dehydrogenase and Fumarate HydrataseGeanelly SALGADO URIOSTEGUINo ratings yet
- 1 s2.0 S0960852410008023 MainDocument9 pages1 s2.0 S0960852410008023 MainGeanelly SALGADO URIOSTEGUINo ratings yet
- 2011 - Preparation of Monolithic CatalystsDocument38 pages2011 - Preparation of Monolithic CatalystsGeanelly SALGADO URIOSTEGUINo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
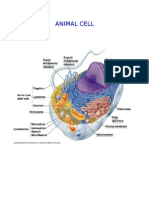
- Animal CellDocument5 pagesAnimal CellNan Fen Jin Sabolarce100% (1)
- Secondary Structure PredictionDocument43 pagesSecondary Structure PredictionPallavi RamaswamyNo ratings yet
- Biology Notes Grade 9Document58 pagesBiology Notes Grade 9Ali Khan100% (2)
- UntitledDocument339 pagesUntitledJOS� FRANCISCO G�MEZ RODR�GUEZNo ratings yet
- Transcription Translation ProtocolDocument13 pagesTranscription Translation ProtocolMrNewbsNo ratings yet
- Fatty Acid Synthesis 2019Document26 pagesFatty Acid Synthesis 2019Yousef KhallafNo ratings yet
- Thesis - Data-Driven Cell Engineering of Chinese Hamster Ovary Cells Through Machine LearningDocument102 pagesThesis - Data-Driven Cell Engineering of Chinese Hamster Ovary Cells Through Machine LearningMaxwell PryceNo ratings yet
- Advanced BIochemistry PPT I SemDocument469 pagesAdvanced BIochemistry PPT I Semkratikaagrawal50No ratings yet
- Chapter 7chromosome Banding and IdentificationDocument18 pagesChapter 7chromosome Banding and IdentificationJed Naziel AvilaNo ratings yet
- Gram StainDocument2 pagesGram StainAltea ArayataNo ratings yet
- Assessment of Hematologic Function and Treatment Modalities Assessment of Hematologic Function and Treatment ModalitiesDocument26 pagesAssessment of Hematologic Function and Treatment Modalities Assessment of Hematologic Function and Treatment ModalitiesEdelweiss Marie CayetanoNo ratings yet
- CAPE BiologyDocument3 pagesCAPE BiologyKashim AliNo ratings yet
- Lesson 7 Cell MembraneDocument3 pagesLesson 7 Cell MembraneThea MillanesNo ratings yet
- Bm7 RemodellingDocument105 pagesBm7 RemodellingabdullahshiddiqadamNo ratings yet
- MIT20 441JF09 Quiz3 SampleDocument4 pagesMIT20 441JF09 Quiz3 SampleGuilherme HenriqueNo ratings yet
- T Cells in Health and DiseaseDocument50 pagesT Cells in Health and Diseaseygilad9139No ratings yet
- BIO 216-Molecular Biology-Sec1-Muhammad Tariq-Shaper Mirza-Syed Shahzad Ul HussanDocument3 pagesBIO 216-Molecular Biology-Sec1-Muhammad Tariq-Shaper Mirza-Syed Shahzad Ul HussanAnonymous sF8ZuiGNo ratings yet
- Micro-Para Answer Key-BLUE PACOPDocument20 pagesMicro-Para Answer Key-BLUE PACOPkelvin raynesNo ratings yet
- Subtopic 4 Carbon BalanceDocument10 pagesSubtopic 4 Carbon BalanceKarla gandaNo ratings yet
- Full Blood CountDocument3 pagesFull Blood CountStephanie SujanaNo ratings yet
- On The Coupling of Mechanics With Bioelectricity and Its Role in MorphogenesisDocument12 pagesOn The Coupling of Mechanics With Bioelectricity and Its Role in MorphogenesisVishvendraNo ratings yet
- Cosmetics: Topical Peptide Treatments With Effective Anti-Aging ResultsDocument14 pagesCosmetics: Topical Peptide Treatments With Effective Anti-Aging ResultsReena BennyNo ratings yet
- Palate Development. Ferguson.Document20 pagesPalate Development. Ferguson.Fabian BarretoNo ratings yet
- Wa0000.Document28 pagesWa0000.mohamed abu mughaisibNo ratings yet
- Ampicillin Vs - CarbenicillinDocument2 pagesAmpicillin Vs - Carbenicillinbeletristicliteratur100% (2)
- Assignment Problem Solving Chapter 2Document2 pagesAssignment Problem Solving Chapter 2Madalyn StarkeyNo ratings yet
- Ms. Rima DessaiDocument46 pagesMs. Rima DessaiSanghaviNo ratings yet
- Bio Exam ReviewerDocument10 pagesBio Exam ReviewerMarlon JonasNo ratings yet
- Food and Chemical Toxicology: SciencedirectDocument27 pagesFood and Chemical Toxicology: SciencedirectDwi PerbriyantiNo ratings yet
- Human Anatomy 7th Edition Martini Test BankDocument32 pagesHuman Anatomy 7th Edition Martini Test BankKerriAdamsdwroi100% (11)