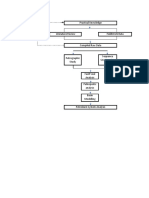
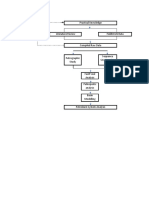
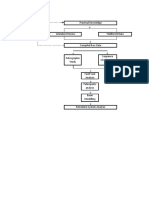
You might also like
- Minimal Residual Disease Testing: Current Innovations and Future DirectionsFrom EverandMinimal Residual Disease Testing: Current Innovations and Future DirectionsTodd E. DruleyNo ratings yet
- Cromatina y ApoptosisDocument11 pagesCromatina y ApoptosisOperacion ConocimientoNo ratings yet
- Cancer Cells Can Exhibit A Sparing FLASH Effect atDocument9 pagesCancer Cells Can Exhibit A Sparing FLASH Effect atMohamed El-AgamyNo ratings yet
- A. Holland: Bioem2015, Inhibition of Human Leukemia Cells in Vitro 1Document6 pagesA. Holland: Bioem2015, Inhibition of Human Leukemia Cells in Vitro 1valorecoragemNo ratings yet
- Sensitive Detection of Tumor Cells Using a Novel 3D-MWCNTs Array CytosensorDocument6 pagesSensitive Detection of Tumor Cells Using a Novel 3D-MWCNTs Array Cytosensorreda rashwanNo ratings yet
- Detection of DNA Damage in Cumulus Cells Using A Chromatin Dispersion AssayDocument10 pagesDetection of DNA Damage in Cumulus Cells Using A Chromatin Dispersion Assay周保亨No ratings yet
- BJR 20190225Document4 pagesBJR 20190225Raul Matute MartinNo ratings yet
- 2015-Oncotarget-Activation of Homologous Recombination DNA Repair in Human Skin Fibroblasts Continuously Exposed To X-Ray RadiationDocument10 pages2015-Oncotarget-Activation of Homologous Recombination DNA Repair in Human Skin Fibroblasts Continuously Exposed To X-Ray Radiation小熊JoeNo ratings yet
- Intracellular Trafficking of Cationic Carbon Dots in Cancer CellDocument13 pagesIntracellular Trafficking of Cationic Carbon Dots in Cancer CellGaby RojasNo ratings yet
- Chromosomal AberrationsDocument20 pagesChromosomal AberrationsArchanaShenoyNo ratings yet
- TMP 610 CDocument6 pagesTMP 610 CFrontiersNo ratings yet
- 02 CGCE Colomuts PublishedDocument6 pages02 CGCE Colomuts PublishedminarikovaluNo ratings yet
- Ultrasound-Mediated DNA Transfer For Bacteria - Yizhi Song Et Al - Nucl Acids Res 2007Document9 pagesUltrasound-Mediated DNA Transfer For Bacteria - Yizhi Song Et Al - Nucl Acids Res 2007Antonio Vázquez MotaNo ratings yet
- tmpDC1A TMPDocument43 pagestmpDC1A TMPFrontiersNo ratings yet
- H2AX in Cancer Cells A Potential Biomarker For Cancer Diagnostics Prediction and RecurrenceDocument6 pagesH2AX in Cancer Cells A Potential Biomarker For Cancer Diagnostics Prediction and RecurrencekikiNo ratings yet
- Dielectric Properties of Breast Carcinoma and The Surrounding TissuesDocument7 pagesDielectric Properties of Breast Carcinoma and The Surrounding TissuesPriya HankareNo ratings yet
- Implications of The Bystander and Abscopal Effects of Radiation TherapyDocument4 pagesImplications of The Bystander and Abscopal Effects of Radiation TherapyDonald ManuainNo ratings yet
- alexiev2014Document5 pagesalexiev2014karen.cobenaNo ratings yet
- Impact of Types of Lymphocyte Chromosomal Aberrations On Human Cancer Risk: Results From Nordic and Italian CohortsDocument6 pagesImpact of Types of Lymphocyte Chromosomal Aberrations On Human Cancer Risk: Results From Nordic and Italian CohortsMuthu KumarNo ratings yet
- 1 s2.0 S0360301620345697 MainDocument13 pages1 s2.0 S0360301620345697 MainXuejun ZhuNo ratings yet
- MMP10 NSCLCDocument9 pagesMMP10 NSCLCbhaskarNo ratings yet
- Electrochimica ActaDocument9 pagesElectrochimica ActagpaivNo ratings yet
- Journal of Photochemistry & Photobiology, B: BiologyDocument8 pagesJournal of Photochemistry & Photobiology, B: BiologyVidal VillanuevaNo ratings yet
- Tumores Mucinosos Del Ovario Con Nodulo Mural Case Report 2020Document10 pagesTumores Mucinosos Del Ovario Con Nodulo Mural Case Report 2020Jairo Lino BNo ratings yet
- Besaratinia 2005Document6 pagesBesaratinia 2005Marcelo DomingosNo ratings yet
- Ol 24 4 13458 PDFDocument9 pagesOl 24 4 13458 PDFCf FungNo ratings yet
- OncologiaDocument16 pagesOncologiaLizbeth Mariscal GaytánNo ratings yet
- Cancer Stem Cells and Signaling Pathways in RadioresistanceDocument16 pagesCancer Stem Cells and Signaling Pathways in RadioresistanceJuan Carlos AlamillaNo ratings yet
- Toussaint Differential ParticipationDocument8 pagesToussaint Differential ParticipationJeffery McDonaldNo ratings yet
- Research Paper On Dna RepairDocument5 pagesResearch Paper On Dna Repairgw060qpy100% (1)
- AugerDocument2 pagesAugerThukscd DlaminiNo ratings yet
- Nanoscale: Accepted ManuscriptDocument14 pagesNanoscale: Accepted ManuscriptSkycancer DukeNo ratings yet
- Original ArticleDocument8 pagesOriginal ArticleMagda MagNo ratings yet
- 3407 FullDocument5 pages3407 FullCyprien YENo ratings yet
- Methods: Wenjuan Liao, Michael A. Mcnutt, Wei-Guo ZhuDocument8 pagesMethods: Wenjuan Liao, Michael A. Mcnutt, Wei-Guo Zhuinoka911No ratings yet
- 09 Light Scattering Measurements of Sub Cellular Structure Providee Noninvasive Early Detection of Chemotherapy-Induced ApoptosisDocument7 pages09 Light Scattering Measurements of Sub Cellular Structure Providee Noninvasive Early Detection of Chemotherapy-Induced ApoptosisLai MattNo ratings yet
- Debucquoy 2006Document6 pagesDebucquoy 2006yalocim666No ratings yet
- Editorials: Cancer, DNA Repair Mechanisms, and Resistance To ChemotherapyDocument3 pagesEditorials: Cancer, DNA Repair Mechanisms, and Resistance To ChemotherapyYuanita AndrianiNo ratings yet
- Electropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsDocument6 pagesElectropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsEspacio UniversitarioNo ratings yet
- Srep 01751Document9 pagesSrep 01751Nadia Maulika Purnama WafiNo ratings yet
- Halo AssayDocument19 pagesHalo AssayMauro Porcel de PeraltaNo ratings yet
- Mechanism and Regulation of Human Non-Homologous Dna End-JoiningDocument9 pagesMechanism and Regulation of Human Non-Homologous Dna End-JoiningAbi Wombat MendiolitaNo ratings yet
- Adewoye2020 Article MultiparameterFlowCytometricDeDocument14 pagesAdewoye2020 Article MultiparameterFlowCytometricDeJose Antonio Cruz CardenasNo ratings yet
- 1 s2.0 S0360301617345066 MainDocument11 pages1 s2.0 S0360301617345066 MainnvkhieuNo ratings yet
- Cytogenetic Follow Up of An Individual After Acc 2021 Mutation Research GeneDocument3 pagesCytogenetic Follow Up of An Individual After Acc 2021 Mutation Research GeneAURORA RENJANI KIRANANo ratings yet
- Doi 10.1016 J.canlet.2007.01.014Document7 pagesDoi 10.1016 J.canlet.2007.01.014aselle kellyNo ratings yet
- PIIS000634951400561XDocument11 pagesPIIS000634951400561XNatalia Lucía Puerta BordaNo ratings yet
- Thesis On Comet AssayDocument4 pagesThesis On Comet Assaymichellebojorqueznorwalk100% (2)
- Experimental Evidence and Model Explanation For Cell Population Characteristics Modification When Applying Sequential Photodynamic TherapyDocument8 pagesExperimental Evidence and Model Explanation For Cell Population Characteristics Modification When Applying Sequential Photodynamic TherapyCamila CamposNo ratings yet
- ijms-22-07504-v2Document35 pagesijms-22-07504-v2usst.almenaraNo ratings yet
- Analysis of Chromosomal Aberrations and H2A - 2021 - Mutation Research GeneticDocument13 pagesAnalysis of Chromosomal Aberrations and H2A - 2021 - Mutation Research GeneticAURORA RENJANI KIRANANo ratings yet
- Colloids and Surfaces B: BiointerfacesDocument10 pagesColloids and Surfaces B: Biointerfacestoyito20No ratings yet
- MC Simulation of Radiation-Induced DNA Double Strand BreaksDocument12 pagesMC Simulation of Radiation-Induced DNA Double Strand BreaksChloe FanningNo ratings yet
- Satb2 2Document6 pagesSatb2 2bela bagus setiawanNo ratings yet
- Experiment and Mechanism Research of SKOV3 Cancer Cell Apoptosis Induced by Nanosecond Pulsed Electric FieldDocument4 pagesExperiment and Mechanism Research of SKOV3 Cancer Cell Apoptosis Induced by Nanosecond Pulsed Electric Fieldjuliogomez008No ratings yet
- Almstrup 2010Document8 pagesAlmstrup 2010Jair BrachoNo ratings yet
- Human Pathology: Case Reports: Kankanamage Malinda Amesh Karasinghe, Kesavan SittampalamDocument5 pagesHuman Pathology: Case Reports: Kankanamage Malinda Amesh Karasinghe, Kesavan SittampalammalindaNo ratings yet
- Human Pathology: Case Reports: Kankanamage Malinda Amesh Karasinghe, Kesavan SittampalamDocument5 pagesHuman Pathology: Case Reports: Kankanamage Malinda Amesh Karasinghe, Kesavan SittampalammalindaNo ratings yet
- Cancer DetectionDocument7 pagesCancer Detectionconference RequirementsNo ratings yet
- The Role of Stereotactic Ablative Body Radiotherapy in Renal Cell CarcinomaDocument10 pagesThe Role of Stereotactic Ablative Body Radiotherapy in Renal Cell CarcinomaRaul Matute MartinNo ratings yet
- Practical Pattern of Surgical Timing of Childhood Cataract in China, A Cross-Sectional Database StudyDocument6 pagesPractical Pattern of Surgical Timing of Childhood Cataract in China, A Cross-Sectional Database StudyAnonymous iInKGkMCNo ratings yet
- Comparison of The Mechanical Properties of The Anterior Lens Capsule in Senile Cataract, Senile Cataract With Trypan Blue Application, and Pseudoexfoliation Syndrome.Document8 pagesComparison of The Mechanical Properties of The Anterior Lens Capsule in Senile Cataract, Senile Cataract With Trypan Blue Application, and Pseudoexfoliation Syndrome.Anonymous iInKGkMCNo ratings yet
- EFFECT OF SHILAJATU RASAYANA IN LETROZOLE INDUCED, Surangi 2018Document2 pagesEFFECT OF SHILAJATU RASAYANA IN LETROZOLE INDUCED, Surangi 2018Anonymous iInKGkMCNo ratings yet
- Estimation of Serum Ascorbic Acid (Vitamin C) in The Age Related (Senile) Cataract: A Case Control StudyDocument5 pagesEstimation of Serum Ascorbic Acid (Vitamin C) in The Age Related (Senile) Cataract: A Case Control StudyAnonymous iInKGkMCNo ratings yet
- Determinan ILODocument6 pagesDeterminan ILOKangJeJaeNo ratings yet
- Bagan Magang PDFDocument1 pageBagan Magang PDFAnonymous iInKGkMCNo ratings yet
- Bagan Magang PDFDocument1 pageBagan Magang PDFAnonymous iInKGkMCNo ratings yet
- Bagan MagangDocument1 pageBagan MagangAnonymous iInKGkMCNo ratings yet
- 3 Molecular and Cellular RadbiologyDocument46 pages3 Molecular and Cellular RadbiologyAngelo RamosNo ratings yet
- Zracenje 1Document22 pagesZracenje 1DejanNo ratings yet
- Radiobiology For The Radiologist 8th Edition Ebook PDFDocument61 pagesRadiobiology For The Radiologist 8th Edition Ebook PDFroy.mula622100% (40)
- Basic radiobiology: the 5 Rs, fractionation, and α/β ratioDocument32 pagesBasic radiobiology: the 5 Rs, fractionation, and α/β ratioAji PatriajatiNo ratings yet
- Radiobiology Essentials for Medical ImagingDocument3 pagesRadiobiology Essentials for Medical ImagingdonnaNo ratings yet
- Radiation Biology OR RadiobiologyDocument36 pagesRadiation Biology OR RadiobiologyVidhyaNo ratings yet
- 4 Radiobiology of Brachytherapy and The Dose-Rate EffectDocument27 pages4 Radiobiology of Brachytherapy and The Dose-Rate EffectDiana MitreaNo ratings yet
- Principles of RadioBio123Document5 pagesPrinciples of RadioBio123riomjNo ratings yet
- Radiation Protection in Medical Radiography 8th Edition Sherer Test BankDocument6 pagesRadiation Protection in Medical Radiography 8th Edition Sherer Test Bankbeatrixkhuyen9rm100% (24)
- RadiotherapyDocument60 pagesRadiotherapyAARYANo ratings yet
- Radiology Exam 2 Notes: Basics of Attenuation, Absorbed Dose, Film ProcessingDocument155 pagesRadiology Exam 2 Notes: Basics of Attenuation, Absorbed Dose, Film ProcessingAprilNelson100% (2)
- A Review of The Use of Radiotherapy in The UKDocument124 pagesA Review of The Use of Radiotherapy in The UKLeandro PolancoNo ratings yet
- Radiation Biology: Presented By: Aarya.H.NairDocument83 pagesRadiation Biology: Presented By: Aarya.H.NairAARYANo ratings yet
- 2 Fundamental Principles of RadiobiologyDocument28 pages2 Fundamental Principles of RadiobiologyAngelo RamosNo ratings yet
- 2 Fundamental Principles of Radiobiology PDFDocument27 pages2 Fundamental Principles of Radiobiology PDFAngelo RamosNo ratings yet
- The Law of Bergonie and Triboundeau A Nice Formula For A First Approximation - Vogin and Foray (2012)Document8 pagesThe Law of Bergonie and Triboundeau A Nice Formula For A First Approximation - Vogin and Foray (2012)DanielNo ratings yet
- Efek Radiasi Dan Proteksi RadiasiDocument56 pagesEfek Radiasi Dan Proteksi RadiasiMargana SitumorangNo ratings yet
- Radiobiology DilshadDocument78 pagesRadiobiology DilshadAJAY K VNo ratings yet
- Complete RadThera Lecture 1Document48 pagesComplete RadThera Lecture 1Soleil SierraNo ratings yet
- Chapter 30 Fundamental Principles of RadiobiologyDocument7 pagesChapter 30 Fundamental Principles of RadiobiologyMegNo ratings yet
- The Main Interaction Process (Es) of Neutrons in Biologic Matter AreDocument19 pagesThe Main Interaction Process (Es) of Neutrons in Biologic Matter ArePshdar QadirNo ratings yet
- Quiz Mock Exam 2 - Ungraded - Requires Respondus Lockdown Browser WebcamDocument60 pagesQuiz Mock Exam 2 - Ungraded - Requires Respondus Lockdown Browser Webcamapi-635954562No ratings yet
- Law of Bergonie and TribondeauDocument7 pagesLaw of Bergonie and TribondeauRuelyn AcedoNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)