You might also like
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5813)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Flames of PerditionDocument16 pagesFlames of Perditiononslowmo100% (4)
- S8Mtiiie-F-10: Matter and Its InteractionsDocument4 pagesS8Mtiiie-F-10: Matter and Its InteractionsMaricris Cagas-PitaoNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Scaffolding GuideDocument143 pagesScaffolding Guideblackwellkid88% (16)
- Hvac For Clean Rooms FacilitiesDocument54 pagesHvac For Clean Rooms FacilitiesAhmed Sherif100% (3)
- Project Management Tools and TechniquesDocument32 pagesProject Management Tools and TechniquesReena Sanehi100% (2)
- Routes of Drug AdministrationDocument36 pagesRoutes of Drug Administrationapule geraldhumbleNo ratings yet
- Module 1 Academic - ScriptDocument10 pagesModule 1 Academic - ScriptArya TrivediNo ratings yet
- Foundations of Inclusive Practice Final ReflectionDocument3 pagesFoundations of Inclusive Practice Final Reflectionapi-533841470No ratings yet
- Extrapolation TablesDocument36 pagesExtrapolation TablesAlex PaicaNo ratings yet
- 1 Millennilas and FilinnialsDocument13 pages1 Millennilas and FilinnialsNix Reora100% (1)
- SEC Presentation (Kelvin) - Singapore Green Labelling Scheme (SGLS)Document53 pagesSEC Presentation (Kelvin) - Singapore Green Labelling Scheme (SGLS)Zachary Yassir GonzalezNo ratings yet
- Simulink StatechartDocument1,408 pagesSimulink StatechartKimberly TaylorNo ratings yet
- The Importance of Learning VocabularyDocument2 pagesThe Importance of Learning VocabularyTrang Nguyễn100% (1)
- Tutorial 2 (207540)Document9 pagesTutorial 2 (207540)Wan Nur AtikahNo ratings yet
- SSDD Lecture Notes A60130 Image Marked TextmarkDocument51 pagesSSDD Lecture Notes A60130 Image Marked TextmarkNikhilesh VarmaNo ratings yet
- Diagnostic Studies For Osteosarcoma: CBC (WBC) ESR Serum Alkaline Phosphatase Alkaline PhosphataseDocument2 pagesDiagnostic Studies For Osteosarcoma: CBC (WBC) ESR Serum Alkaline Phosphatase Alkaline PhosphatasekyawNo ratings yet
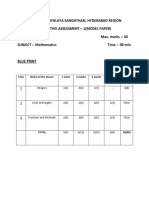
- Cbse Sample Papers For Class 7 Maths Fa4 PDFDocument5 pagesCbse Sample Papers For Class 7 Maths Fa4 PDFGirish LalaNo ratings yet
- Report 1668349748488Document86 pagesReport 1668349748488Victor TabuncicNo ratings yet
- NagarjunaDocument4 pagesNagarjunaCLG100% (1)
- A. Dunn, The Exploitation and Control of Woodland and Scrubland in The Byzantine WorldDocument64 pagesA. Dunn, The Exploitation and Control of Woodland and Scrubland in The Byzantine WorldSidiropoulos DimitriosNo ratings yet
- Full Download Solution Manual For Introduction To Derivatives and Risk Management 9th Edition Don M Chance Robert Brooks PDF Full ChapterDocument20 pagesFull Download Solution Manual For Introduction To Derivatives and Risk Management 9th Edition Don M Chance Robert Brooks PDF Full Chapterdiclinicauroravsl9100% (20)
- Ultra-Slim, 4.2 MM Diameter Videoscope With 2.0 MM Instrument Channel Enables Better Access To Peripheral AreasDocument2 pagesUltra-Slim, 4.2 MM Diameter Videoscope With 2.0 MM Instrument Channel Enables Better Access To Peripheral AreaskrsrinivasarajuNo ratings yet
- RA-013205 - MASTER PLUMBER - Manila - 2-2022Document112 pagesRA-013205 - MASTER PLUMBER - Manila - 2-2022Rainier VillanuevaNo ratings yet
- Solar Parks: Accelerating The Growth of Solar Power in IndiaDocument5 pagesSolar Parks: Accelerating The Growth of Solar Power in IndiaNiket DalalNo ratings yet
- Physicochemical Properties and Sensory Attributes of New Formulations of Dry-Fermented SausagesDocument5 pagesPhysicochemical Properties and Sensory Attributes of New Formulations of Dry-Fermented SausagesShermsooddeen Asheer EvraheemNo ratings yet
- Psalms, Hymns, and Spiritual Songs: The Master Musician's MelodiesDocument4 pagesPsalms, Hymns, and Spiritual Songs: The Master Musician's MelodiesAlexey LevchukNo ratings yet
- Soft Computing Fundamentals and Applications EbookDocument4 pagesSoft Computing Fundamentals and Applications EbookGrammarly AccountNo ratings yet
- FYP Report MDocument33 pagesFYP Report MSumaira SaifNo ratings yet
- Enabling SSL For Enterprise ApplicationsDocument22 pagesEnabling SSL For Enterprise ApplicationsSaket ShrivastavNo ratings yet
- Volvo V90 Cross Country, La Scheda TecnicaDocument3 pagesVolvo V90 Cross Country, La Scheda TecnicaBlogo MotoriNo ratings yet