You might also like
- Basic Neurochemistry: Principles of Molecular, Cellular, and Medical NeurobiologyFrom EverandBasic Neurochemistry: Principles of Molecular, Cellular, and Medical NeurobiologyNo ratings yet
- Proyecto de Tesis - Nicole SanguinettiDocument26 pagesProyecto de Tesis - Nicole SanguinettiPau Fer Mujica CNo ratings yet
- Neurotransmisores Del Sistema LimbicoDocument9 pagesNeurotransmisores Del Sistema LimbicoEli CastilloNo ratings yet
- Pi Is 0006349520314788Document2 pagesPi Is 0006349520314788Sena 1452No ratings yet
- Levin - Godukhin - 2017 - Modulating Effect of Cytokines On Mechanisms of Synaptic Plasticity in The BrainDocument11 pagesLevin - Godukhin - 2017 - Modulating Effect of Cytokines On Mechanisms of Synaptic Plasticity in The Brainkeon.arbabi.altNo ratings yet
- GABA Depolarizes Immature NeuronsDocument13 pagesGABA Depolarizes Immature NeuronsDaniel ChaguenNo ratings yet
- Annurev - Castillo PDocument27 pagesAnnurev - Castillo PSebastián GallegosNo ratings yet
- Artigo Bioquímica 1Document9 pagesArtigo Bioquímica 1MCapitolinaNo ratings yet
- ESSAY BREZAN SIMON Biology Lectures 2006 - Neuroscience Summer School at FORTH Institute: Onassis Foundation Award - StipendDocument16 pagesESSAY BREZAN SIMON Biology Lectures 2006 - Neuroscience Summer School at FORTH Institute: Onassis Foundation Award - Stipendsimon brežanNo ratings yet
- AS Bases Celulares DE LAS Oscilaciones Neuronales: UmmaryDocument8 pagesAS Bases Celulares DE LAS Oscilaciones Neuronales: Ummarydavid alejandro pinzon sanchezNo ratings yet
- Neuromodulation of Hippocampal Synaptic Plasticity, Learning, and Memory by NoradrenalineDocument17 pagesNeuromodulation of Hippocampal Synaptic Plasticity, Learning, and Memory by NoradrenalinecarbonloveNo ratings yet
- 5 GabaDocument11 pages5 GabaJose GiovannyNo ratings yet
- Margarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisDocument12 pagesMargarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisJmasnNo ratings yet
- N - I. A - P: Eurotransmisores DEL Sistema Límbico Mígdala Rimera ParteDocument6 pagesN - I. A - P: Eurotransmisores DEL Sistema Límbico Mígdala Rimera ParteghgfchmNo ratings yet
- Diacylglycerol Lipase A ManipulationDocument9 pagesDiacylglycerol Lipase A ManipulationKis AlexandraNo ratings yet
- BDNF en PCKDocument8 pagesBDNF en PCKJesusNo ratings yet
- Introduction To Neural Plasticity MechanismDocument9 pagesIntroduction To Neural Plasticity MechanismAlain CésarNo ratings yet
- Tepper and Bolam, 2004 Curr Opin NeurobiolDocument8 pagesTepper and Bolam, 2004 Curr Opin NeurobiolSarah Miryam CoffanoNo ratings yet
- NeuropharmacologyDocument20 pagesNeuropharmacologysajveen100% (1)
- Role of GABAergic Inhibition in HippocamDocument7 pagesRole of GABAergic Inhibition in HippocamdagushNo ratings yet
- Author's Accepted Manuscript: Biological PsychiatryDocument31 pagesAuthor's Accepted Manuscript: Biological PsychiatryGustav RSRNo ratings yet
- PNAS 2001 West 11024 31Document8 pagesPNAS 2001 West 11024 31Vehpi YILDIRIMNo ratings yet
- Limbic and Cortical Information Processing in The Nucleus AccumbensDocument7 pagesLimbic and Cortical Information Processing in The Nucleus AccumbensEszter SzerencsiNo ratings yet
- Brain Neurosteroids Are 4th Generation Neuromessengers in The Brain: Cell Biophysical Analysis of Steroid Signal TransductionDocument30 pagesBrain Neurosteroids Are 4th Generation Neuromessengers in The Brain: Cell Biophysical Analysis of Steroid Signal TransductionLonkesNo ratings yet
- Clock GenesDocument16 pagesClock GenesPRASHANT SOLANKINo ratings yet
- Reward Circuitry in Addiction: ReviewDocument11 pagesReward Circuitry in Addiction: ReviewagusalienNo ratings yet
- Role of Lipid Rafts and The Underlying - Chapter 64Document13 pagesRole of Lipid Rafts and The Underlying - Chapter 64Christina MountakiNo ratings yet
- Pathophysiology of EpilepsyDocument16 pagesPathophysiology of EpilepsyYexalen Zazil100% (1)
- Symposium 2023 AbstractsDocument21 pagesSymposium 2023 Abstractsmail2umamaheswari15No ratings yet
- Ni Hms 21239Document9 pagesNi Hms 21239Ionut NeagaNo ratings yet
- Ubiquitin, The Proteasome and Protein Degradation in Neuronal Function and DysfunctionDocument13 pagesUbiquitin, The Proteasome and Protein Degradation in Neuronal Function and DysfunctionalbertoNo ratings yet
- Daniel O. Kellett Et Al - Memory Consolidation in The Cerebellar CortexDocument13 pagesDaniel O. Kellett Et Al - Memory Consolidation in The Cerebellar CortexCortate15gNo ratings yet
- Epilepsy and Seizures: PathophysiologyDocument11 pagesEpilepsy and Seizures: PathophysiologyAbrizan HassanNo ratings yet
- Jurnal 5 CDocument9 pagesJurnal 5 CCintya RambuNo ratings yet
- Aims and Significance: Protti Sesqui R & D - 1Document5 pagesAims and Significance: Protti Sesqui R & D - 1Jackson ChenNo ratings yet
- Clock GenesDocument8 pagesClock GenesPopsNo ratings yet
- Clase 5 - YamadaDocument11 pagesClase 5 - YamadaCarlos Francisco Rincon LozadaNo ratings yet
- Cotransmission: Geoffrey BurnstockDocument6 pagesCotransmission: Geoffrey BurnstockSuryasumanthNo ratings yet
- Cannabis Paper 7Document13 pagesCannabis Paper 7Carlos Eduardo Jorquera GonzálezNo ratings yet
- Ni Hms 262290Document8 pagesNi Hms 262290Ignacio MazzitelliNo ratings yet
- Neuromodulation of Prefrontal Cortex Cognitive Function in Primates - The Powerful Roles of Monoamines and AcetylcholineDocument20 pagesNeuromodulation of Prefrontal Cortex Cognitive Function in Primates - The Powerful Roles of Monoamines and AcetylcholineDouglas DiasNo ratings yet
- Protein Kinase-Based Neural Signaling Pathways For Ginsenosides: A Retrospective ReviewDocument6 pagesProtein Kinase-Based Neural Signaling Pathways For Ginsenosides: A Retrospective Reviewnotapernota101No ratings yet
- Muscarinic Receptors: Their Roles in Disorders of The Central Nervous System and Potential As Therapeutic TargetsDocument11 pagesMuscarinic Receptors: Their Roles in Disorders of The Central Nervous System and Potential As Therapeutic TargetsDavide MandelliNo ratings yet
- Neuro PlasticityDocument67 pagesNeuro Plasticitydrkadiyala2100% (1)
- Synaptic TransmissionDocument17 pagesSynaptic TransmissionbnmjgcNo ratings yet
- Introduction To CNS PharmacologyDocument49 pagesIntroduction To CNS Pharmacologymatchees-gone rogueNo ratings yet
- Splice Variants of The B-Site APP-cleaving Enzyme BACE1 in Human Brain and PancreasDocument8 pagesSplice Variants of The B-Site APP-cleaving Enzyme BACE1 in Human Brain and PancreasAlix AliNo ratings yet
- Acsády Et Al. - 2000 - Nerve Growth Factor But Not Neurotrophin-3 Is Synthesized by Hippocampal GABAergic Neurons That Project To The MeDocument9 pagesAcsády Et Al. - 2000 - Nerve Growth Factor But Not Neurotrophin-3 Is Synthesized by Hippocampal GABAergic Neurons That Project To The MeGabriel HerreraNo ratings yet
- Dr. Shamsher Singh and SunpreetDocument17 pagesDr. Shamsher Singh and SunpreetSUNPREET KAURNo ratings yet
- BMC NeuroscienceDocument7 pagesBMC NeuroscienceDmitry DiykovNo ratings yet
- Velisek 2017 Puritan S View of The Menage A TroisDocument3 pagesVelisek 2017 Puritan S View of The Menage A TroissilenceindigoNo ratings yet
- 2009 - Shaw, FilbertDocument10 pages2009 - Shaw, FilbertAfranio Cogo DestefaniNo ratings yet
- CANal de GABA en BNST en InglesDocument18 pagesCANal de GABA en BNST en InglesPinPunkNo ratings yet
- Brozoski 2005Document6 pagesBrozoski 2005Caesar Catalin CaratasuNo ratings yet
- Cocaine-mediated circadian reprogramming in the striatum through dopamine D2R and Ppar γ activationDocument14 pagesCocaine-mediated circadian reprogramming in the striatum through dopamine D2R and Ppar γ activationGabriele SantosNo ratings yet
- (Kuliah 3) Mekanisme Akupunktur Medik Sari18Document65 pages(Kuliah 3) Mekanisme Akupunktur Medik Sari18callNo ratings yet
- Epc 13771Document3 pagesEpc 13771Van Basten ArdiansyachNo ratings yet
- Reviews: Overview of Pdes and Their RegulationDocument19 pagesReviews: Overview of Pdes and Their RegulationAli HassanNo ratings yet
- Neurobiology of Learning and MemoryDocument7 pagesNeurobiology of Learning and MemoryNothingNo ratings yet
- Romantic Love An fMRI Study of Aneural Mechanism For Mate ChoiceDocument5 pagesRomantic Love An fMRI Study of Aneural Mechanism For Mate ChoiceArnt van HeldenNo ratings yet
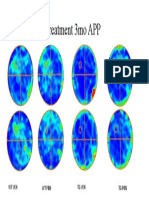
- MWM Pre Treatment 18 MO APP: WT1 WT2 TG1 TG2Document1 pageMWM Pre Treatment 18 MO APP: WT1 WT2 TG1 TG2Pau Fer Mujica CNo ratings yet
- MWM Post Treatment 3mo APP: WT Veh WT PBN TG Veh TG PBNDocument1 pageMWM Post Treatment 3mo APP: WT Veh WT PBN TG Veh TG PBNPau Fer Mujica CNo ratings yet
- MWM Post Treatment 18mo APP: WT Veh WT PBN TG Veh TG PBNDocument1 pageMWM Post Treatment 18mo APP: WT Veh WT PBN TG Veh TG PBNPau Fer Mujica CNo ratings yet
- Weight Clips (4) : Viral InjectionDocument3 pagesWeight Clips (4) : Viral InjectionPau Fer Mujica CNo ratings yet
- NOR Post Treatment 18mo APP: WT Veh TG Veh TG PBN WT PBNDocument1 pageNOR Post Treatment 18mo APP: WT Veh TG Veh TG PBN WT PBNPau Fer Mujica CNo ratings yet
- MWM Pre Treatment 12mo APP: WT Veh TG VehDocument1 pageMWM Pre Treatment 12mo APP: WT Veh TG VehPau Fer Mujica CNo ratings yet
- Genotyping IN - Cre OriginalDocument1 pageGenotyping IN - Cre OriginalPau Fer Mujica CNo ratings yet
- 15.2.1. A, P, B R: Nesthesia Erfusion AND Rain EmovalDocument6 pages15.2.1. A, P, B R: Nesthesia Erfusion AND Rain EmovalPau Fer Mujica CNo ratings yet
- Boletín Mensual Del CINVDocument7 pagesBoletín Mensual Del CINVPau Fer Mujica CNo ratings yet
- Warren McCulloch - A Historical Introduction To The Postulational Foundations of Experimental EpistemologyDocument8 pagesWarren McCulloch - A Historical Introduction To The Postulational Foundations of Experimental EpistemologylazarosirdanielNo ratings yet
- Essentials of Anatomy and Physiology 6th Edition Martini Solutions ManualDocument12 pagesEssentials of Anatomy and Physiology 6th Edition Martini Solutions Manualscarletba4cc100% (34)
- Markscheme Unit5 (6BI05) June2013Document23 pagesMarkscheme Unit5 (6BI05) June2013Mahiat Miftah ShowkatNo ratings yet
- Coordination and Control (IGCSE and O Level Biology 2023-2025)Document65 pagesCoordination and Control (IGCSE and O Level Biology 2023-2025)Tashwaar TamimNo ratings yet
- 500 Words Essay ExampleDocument44 pages500 Words Essay Exampleafhbgufyf100% (2)
- Neurons and Synapses: Essential Idea: Neurons Transmit The Message, Synapses Modulate The MessageDocument26 pagesNeurons and Synapses: Essential Idea: Neurons Transmit The Message, Synapses Modulate The MessageSeanLaiNo ratings yet
- Sensation and Perception 2nd Edition Test BankDocument28 pagesSensation and Perception 2nd Edition Test BanktestbanklooNo ratings yet
- CH 48Document11 pagesCH 48randomtamaNo ratings yet
- MID YEAR Assessment - SCIENCE10Document5 pagesMID YEAR Assessment - SCIENCE10anelou guingueNo ratings yet
- The A To Z of Peripheral NervesDocument230 pagesThe A To Z of Peripheral NervesAspenPharma95% (19)
- Multiple-Choice Questions: For All Questions, Select The Single Best Answer Unless Otherwise DirectedDocument67 pagesMultiple-Choice Questions: For All Questions, Select The Single Best Answer Unless Otherwise DirectedShah NawazNo ratings yet
- Brianbee Sample Questions 1 PDFDocument2 pagesBrianbee Sample Questions 1 PDFLigeia100% (1)
- PYC3703 Study Notes 2013 SummeryDocument60 pagesPYC3703 Study Notes 2013 SummeryErdanx Sanchez100% (1)
- Gross Anatomy MNTSDocument354 pagesGross Anatomy MNTSSolomon Seth SallforsNo ratings yet
- 2016 Brain Bee Competitions Questions - DoDocument10 pages2016 Brain Bee Competitions Questions - DoughNo ratings yet
- Intro2007 Midterm PDFDocument22 pagesIntro2007 Midterm PDFRauMuốngXàoTỏiNo ratings yet
- Kriya Breath PDFDocument1 pageKriya Breath PDFCarmenNo ratings yet
- 2013 - Tibau Valencia Soriano - Frontiers in Neural Circuites - Neuronal Net Properties and Ca ImagingDocument16 pages2013 - Tibau Valencia Soriano - Frontiers in Neural Circuites - Neuronal Net Properties and Ca ImagingmolmedchemNo ratings yet
- Synaptic TransmissionDocument35 pagesSynaptic TransmissiondehaNo ratings yet
- Soft Computing MCQDocument12 pagesSoft Computing MCQANUJ85% (13)
- TMP 26 FDDocument64 pagesTMP 26 FDFrontiersNo ratings yet
- Shull, D. (2003) - Neurobiology of Repetition Compulsion. Annals of Modern Psychoanalysis, 2 (1), 21-46.Document26 pagesShull, D. (2003) - Neurobiology of Repetition Compulsion. Annals of Modern Psychoanalysis, 2 (1), 21-46.Shawn A. Wygant100% (1)
- OCR A Level Biology A SB2 AnswersDocument58 pagesOCR A Level Biology A SB2 AnswersHajin Dho100% (1)
- ch07 - The Nervous SystemDocument25 pagesch07 - The Nervous SystemEbtihal AlharthiNo ratings yet
- Artificial Neural NetworksDocument31 pagesArtificial Neural NetworksDeepa ParasarNo ratings yet
- Child Development A Cultural Approach 2nd Edition Arnett Test Bank 1Document117 pagesChild Development A Cultural Approach 2nd Edition Arnett Test Bank 1joshua100% (36)
- The Nervous SystemDocument57 pagesThe Nervous SystemGeelin Crizza AlbisNo ratings yet
- CNS NeurotransmittersDocument30 pagesCNS Neurotransmittersdevanshi_582262183100% (1)
- Anatomy and PhysiologyDocument8 pagesAnatomy and Physiologymhel03_chickmagnetNo ratings yet
- Science G 6 DappDocument6 pagesScience G 6 DappChester Austin Reese Maslog Jr.No ratings yet