You might also like
- Molecular Biology Techniques: A Classroom Laboratory ManualFrom EverandMolecular Biology Techniques: A Classroom Laboratory ManualNo ratings yet
- Early Diagnosis of A Mexican Variant of Papaya Meleira Virus (Pmev-Mx) by RT-PCRDocument10 pagesEarly Diagnosis of A Mexican Variant of Papaya Meleira Virus (Pmev-Mx) by RT-PCRWendy SerraNo ratings yet
- Cristina Romero RodríguezDocument7 pagesCristina Romero Rodríguezsouidi.zNo ratings yet
- The Complete Sequence of A Papaya Ringspot Virus (PRSV) Isolate From West Bengal, India Infecting Papaya and Study of Genetic VariationDocument8 pagesThe Complete Sequence of A Papaya Ringspot Virus (PRSV) Isolate From West Bengal, India Infecting Papaya and Study of Genetic VariationSyarifah TasharaNo ratings yet
- Cladistic Relationships Among The Pleurotus Ostreatus Complex, The Pleurotus Pulmonarius Complex, and Pleurotus Eryngii Based On The Mitochondrial Small Subunit Ribosomal DNA Sequence AnalysisDocument6 pagesCladistic Relationships Among The Pleurotus Ostreatus Complex, The Pleurotus Pulmonarius Complex, and Pleurotus Eryngii Based On The Mitochondrial Small Subunit Ribosomal DNA Sequence AnalysisAlejandro Murillo V.No ratings yet
- 5-Microbiology Resource Announcements - 2019 - Val-Calvo J Et, AlDocument2 pages5-Microbiology Resource Announcements - 2019 - Val-Calvo J Et, AlAntonNo ratings yet
- Jurnal Kel 6Document8 pagesJurnal Kel 6Rahmy KartikaNo ratings yet
- Pengembangan Penanda Molekuler Untuk Deteksi Phytophthora Palmivora Pada Tanaman KakaoDocument10 pagesPengembangan Penanda Molekuler Untuk Deteksi Phytophthora Palmivora Pada Tanaman KakaoAnita GustinaNo ratings yet
- Unknown - 2010 - Book Announcements Book ShelfDocument209 pagesUnknown - 2010 - Book Announcements Book Shelfmarcos_de_carvalhoNo ratings yet
- Rickettsia en CubaDocument4 pagesRickettsia en CubaDaniel Alfredo Galindo ChoqueNo ratings yet
- Snounou, G. Et Al. High Sensitivity of Detection of Human Malaria Parasites by The Use of Nested Polymerase Chain ReactionDocument7 pagesSnounou, G. Et Al. High Sensitivity of Detection of Human Malaria Parasites by The Use of Nested Polymerase Chain ReactionJorge Mori MarinNo ratings yet
- Genetic Characterization of Amazonian Bovine Papillomavirus Reveals The Existence of Four New Putative TypesDocument8 pagesGenetic Characterization of Amazonian Bovine Papillomavirus Reveals The Existence of Four New Putative TypespafspontesNo ratings yet
- Rna ThesisDocument6 pagesRna Thesiscarlabenningtonnaperville100% (2)
- PD 90 1004Document8 pagesPD 90 1004bayamcrispyyNo ratings yet
- Molecular Characterisation of A Novel and Highly Divergent Passerine Adenovirus 1Document16 pagesMolecular Characterisation of A Novel and Highly Divergent Passerine Adenovirus 1Abdul jNo ratings yet
- Naseem2016 PDFDocument7 pagesNaseem2016 PDFRXMacBlend EswarJRNo ratings yet
- Morfologi Tumbuhan Ketumbar Wahyuni Et Al PDFDocument11 pagesMorfologi Tumbuhan Ketumbar Wahyuni Et Al PDFals alamNo ratings yet
- ISSR Markers As A Tool For The Assessment of Genetic Diversity in Passi OraDocument16 pagesISSR Markers As A Tool For The Assessment of Genetic Diversity in Passi OraAntonia MartinezNo ratings yet
- Molecular TechniquesDocument4 pagesMolecular TechniquesMulatuNo ratings yet
- 11492-Article Text-15660-1-10-20200916Document6 pages11492-Article Text-15660-1-10-20200916bastian herreraNo ratings yet
- Phylogenetic Relationships Amongst 10 Durio Species Based On PCR-RFLP Analysis of Two Chloroplast GenesDocument8 pagesPhylogenetic Relationships Amongst 10 Durio Species Based On PCR-RFLP Analysis of Two Chloroplast GenesFauzanNo ratings yet
- Molecular Diversity and Genetic Structure of Guineagrass (Panicum Maximum Jacq.), A Tropical Pasture GrassDocument19 pagesMolecular Diversity and Genetic Structure of Guineagrass (Panicum Maximum Jacq.), A Tropical Pasture GrassnarayananNo ratings yet
- Grupo 6. Perez Et Al. 2016Document14 pagesGrupo 6. Perez Et Al. 2016Khebyn Esquía AguilarNo ratings yet
- Diversity and Biotechnological Potential of Culturable Bacteria From Brazilian Mangrove SedimentDocument8 pagesDiversity and Biotechnological Potential of Culturable Bacteria From Brazilian Mangrove SedimentMuhammad Wildan GunaryaNo ratings yet
- First Report of Amblyomma Sp. Collected From Varanus SalvatorDocument6 pagesFirst Report of Amblyomma Sp. Collected From Varanus Salvatorbambang awanNo ratings yet
- 1 s2.0 S1413867012001948 MainkDocument7 pages1 s2.0 S1413867012001948 MainkJamilah LucenaNo ratings yet
- Benh VirusDocument7 pagesBenh VirusThu Hien NguyenNo ratings yet
- Bell PepperDocument1 pageBell Pepperkomaltahir2021No ratings yet
- First Detection of Adenovirus in The Vampire Bat (Desmodus Rotundus) in BrazilDocument4 pagesFirst Detection of Adenovirus in The Vampire Bat (Desmodus Rotundus) in BrazilRenata BudaszewskiNo ratings yet
- 2010 (Dib Et Al.) Novel Linear Megaplasmid From Brevibacterium Sp. Isolated From Extreme EnvironmentDocument5 pages2010 (Dib Et Al.) Novel Linear Megaplasmid From Brevibacterium Sp. Isolated From Extreme EnvironmentDaniel AlonsoNo ratings yet
- Accepted Manuscript: Mammarenavirus) Naturally InfectingDocument23 pagesAccepted Manuscript: Mammarenavirus) Naturally InfectingLúcia ReisNo ratings yet
- RAPD-PCR Based Marker Approach For The Genetic Differentiation of Two Species of Cockroach (Order-Dictyoptera)Document6 pagesRAPD-PCR Based Marker Approach For The Genetic Differentiation of Two Species of Cockroach (Order-Dictyoptera)LifedavidNo ratings yet
- Identifying Paracoccidioides Phylogenetic Species by PCR-RFLP of The Alpha-Tubulin GeneDocument8 pagesIdentifying Paracoccidioides Phylogenetic Species by PCR-RFLP of The Alpha-Tubulin GeneKirana agist wangsa putriNo ratings yet
- s00705 021 05348 9 PDFDocument5 pagess00705 021 05348 9 PDFmesele tilahunNo ratings yet
- Discusión Art. OmicasDocument6 pagesDiscusión Art. OmicasARIZBETH GUADALUPE VILLANUEVA FLORESNo ratings yet
- Infectious Hypodermal and Hematopoietic Necrosis Virus From Bra 2014 Virus RDocument11 pagesInfectious Hypodermal and Hematopoietic Necrosis Virus From Bra 2014 Virus RTanIsmNo ratings yet
- Molecular Characterization of Wild MushroomDocument5 pagesMolecular Characterization of Wild Mushroommanojtbgri5793No ratings yet
- Raman Spectroscopy Enables Non-Invasive Identification of Peanut Genotypes and Value-Added TraitsDocument10 pagesRaman Spectroscopy Enables Non-Invasive Identification of Peanut Genotypes and Value-Added TraitsFábio UcellaNo ratings yet
- González2003 Article AComparativeSequenceAnalysisToDocument29 pagesGonzález2003 Article AComparativeSequenceAnalysisToDanilo RosaNo ratings yet
- Tracing The Origins of The Pituitary Adenylate-Cyclase Activating Polypeptide (PACAP)Document19 pagesTracing The Origins of The Pituitary Adenylate-Cyclase Activating Polypeptide (PACAP)ManoloGarridoNo ratings yet
- Arabidopsis Thaliana Research PaperDocument5 pagesArabidopsis Thaliana Research Paperafnhgssontbxkd100% (1)
- Characterization and Molecular Methods For Detection of A Novel Spiroplasma Pathogenic To Penaeus VannameiDocument11 pagesCharacterization and Molecular Methods For Detection of A Novel Spiroplasma Pathogenic To Penaeus Vannameikiura_escalanteNo ratings yet
- International Journal of Microbiology Research: January 2013Document7 pagesInternational Journal of Microbiology Research: January 2013DDRNo ratings yet
- Plant Breeding - 2021 - Zhang - Development of Codominant SCAR Markers To Detect The Pto Tm22 I3 and Sw5 Genes in Tomato PDFDocument7 pagesPlant Breeding - 2021 - Zhang - Development of Codominant SCAR Markers To Detect The Pto Tm22 I3 and Sw5 Genes in Tomato PDFKERTÉSZMÉRNÖK SZIEMKKNo ratings yet
- Molecular Detection and Phylogenetic Analysis of Lumpy Skin Disease Virus From Clinically Infected Cattle of Andhra PradeshDocument8 pagesMolecular Detection and Phylogenetic Analysis of Lumpy Skin Disease Virus From Clinically Infected Cattle of Andhra PradeshSaikumar GangitlaNo ratings yet
- Jannotti Passos, 2010Document8 pagesJannotti Passos, 2010xicoalexandreNo ratings yet
- Genome Sequences of Bacillus Sporothermodurans Strains Isolated From Ultra-High-Temperature MilkDocument3 pagesGenome Sequences of Bacillus Sporothermodurans Strains Isolated From Ultra-High-Temperature Milkfelipeloz087069No ratings yet
- Improved Detection of Citrus Psorosis Virus and Coat ProteinDerived Transgenes in Citrus Plants Comparison Between RTQPCR and TASELISADocument11 pagesImproved Detection of Citrus Psorosis Virus and Coat ProteinDerived Transgenes in Citrus Plants Comparison Between RTQPCR and TASELISAMayra GameroNo ratings yet
- Diversity of Aspergillus isolates and selection of an isolate with high β-fructofuranosidase activity that is native to the Peruvian coastDocument9 pagesDiversity of Aspergillus isolates and selection of an isolate with high β-fructofuranosidase activity that is native to the Peruvian coastJuan CisnerosNo ratings yet
- Artigo - 6 - Cardinali-Rezende - Genome Announc.-2015-Cardinali-Rezende PDFDocument2 pagesArtigo - 6 - Cardinali-Rezende - Genome Announc.-2015-Cardinali-Rezende PDFJuliana Cardinali RezendeNo ratings yet
- Seminar Michael Christian 05 SignedDocument11 pagesSeminar Michael Christian 05 SignedLufia CeruleanNo ratings yet
- ECBF4A762913Document7 pagesECBF4A762913João Gilberto Alves VillelaNo ratings yet
- First Confirmed Report of Basal Rot of Chamaelucium Uncinatum Caused by Phytophthora Nicotianae in ArgentinaDocument3 pagesFirst Confirmed Report of Basal Rot of Chamaelucium Uncinatum Caused by Phytophthora Nicotianae in ArgentinaDaniel VilasecoNo ratings yet
- Molecular Identification Methods of Fish Species: Reassessment and Possible ApplicationsDocument29 pagesMolecular Identification Methods of Fish Species: Reassessment and Possible ApplicationsGlennice Joie SerradaNo ratings yet
- CitriDocument4 pagesCitriAnonymous RkI7MG6geNo ratings yet
- Contracaecum Larvae: Morphological and Morphometric Retrospective Analysis, Biogeography and Zoonotic Risk in The AmazonDocument22 pagesContracaecum Larvae: Morphological and Morphometric Retrospective Analysis, Biogeography and Zoonotic Risk in The AmazonDavid CultureNo ratings yet
- A Novel Use of Infrared SpectrosDocument12 pagesA Novel Use of Infrared Spectroseergenekonn1No ratings yet
- Enfermedad Periodontal ArticuloDocument14 pagesEnfermedad Periodontal ArticuloRauMoliNo ratings yet
- PDA-Global Oct 2011 Poster MR2Document1 pagePDA-Global Oct 2011 Poster MR2sblazej7118No ratings yet
- 1 s2.0 S0378874114006722 MainDocument8 pages1 s2.0 S0378874114006722 MainJan Clarence DaroyNo ratings yet
- Network Analysis of The Papaya Orchard Virome From Two Agroecological Regions of Chiapas, MexicoDocument17 pagesNetwork Analysis of The Papaya Orchard Virome From Two Agroecological Regions of Chiapas, MexicoWendy SerraNo ratings yet
- An Overview of Global Papaya ProductionDocument6 pagesAn Overview of Global Papaya ProductionWendy SerraNo ratings yet
- Identification of Phytoplasma in Six Fruit Crops in IndiaDocument10 pagesIdentification of Phytoplasma in Six Fruit Crops in IndiaWendy SerraNo ratings yet
- Abreu 2 Review PSDDocument18 pagesAbreu 2 Review PSDWendy SerraNo ratings yet
- 10.1007@s10658 020 02019 4Document7 pages10.1007@s10658 020 02019 4Wendy SerraNo ratings yet
- Host Weed Species Range of Meloidogyne Ethiopica Whitehead (Tylenchida: Meloidogynidae) Found in BrazilDocument7 pagesHost Weed Species Range of Meloidogyne Ethiopica Whitehead (Tylenchida: Meloidogynidae) Found in BrazilWendy SerraNo ratings yet
- 10.1007@s10658 020 01962 6Document7 pages10.1007@s10658 020 01962 6Wendy SerraNo ratings yet
- Cercospora Cf. Nicotianae Is A Causal Agent of Cercospora Leaf Blight of SoybeanDocument5 pagesCercospora Cf. Nicotianae Is A Causal Agent of Cercospora Leaf Blight of SoybeanWendy SerraNo ratings yet
- Ron Kangas - IoanDocument11 pagesRon Kangas - IoanBogdan SoptereanNo ratings yet
- Study 107 - The Doctrine of Salvation - Part 8Document2 pagesStudy 107 - The Doctrine of Salvation - Part 8Jason MyersNo ratings yet
- Wilcoxon Matched Pairs Signed Rank TestDocument3 pagesWilcoxon Matched Pairs Signed Rank TestDawn Ilish Nicole DiezNo ratings yet
- Agnes de MilleDocument3 pagesAgnes de MilleMarie-Maxence De RouckNo ratings yet
- Companyprofil E: Erfanconstructionsolut IonDocument14 pagesCompanyprofil E: Erfanconstructionsolut IonNurin AleesyaNo ratings yet
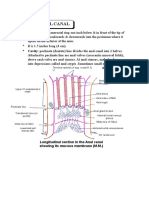
- Anatomy Anal CanalDocument14 pagesAnatomy Anal CanalBela Ronaldoe100% (1)
- Fss Presentation Slide GoDocument13 pagesFss Presentation Slide GoReinoso GreiskaNo ratings yet
- ISBN Safe Work Method Statements 2022 03Document8 pagesISBN Safe Work Method Statements 2022 03Tamo Kim ChowNo ratings yet
- Soft Ground Improvement Using Electro-Osmosis.Document6 pagesSoft Ground Improvement Using Electro-Osmosis.Vincent Ling M SNo ratings yet
- ReadmeDocument3 pagesReadmedhgdhdjhsNo ratings yet
- Functional DesignDocument17 pagesFunctional DesignRajivSharmaNo ratings yet
- Task 3 - LPDocument21 pagesTask 3 - LPTan S YeeNo ratings yet
- Leigh Shawntel J. Nitro Bsmt-1A Biostatistics Quiz No. 3Document6 pagesLeigh Shawntel J. Nitro Bsmt-1A Biostatistics Quiz No. 3Lue SolesNo ratings yet
- FMC Derive Price Action GuideDocument50 pagesFMC Derive Price Action GuideTafara MichaelNo ratings yet
- Ito Na Talaga Yung FinalDocument22 pagesIto Na Talaga Yung FinalJonas Gian Miguel MadarangNo ratings yet
- 04 - Fetch Decode Execute Cycle PDFDocument3 pages04 - Fetch Decode Execute Cycle PDFShaun HaxaelNo ratings yet
- Unit 7: Anthropology: Q2e Listening & Speaking 4: Audio ScriptDocument6 pagesUnit 7: Anthropology: Q2e Listening & Speaking 4: Audio ScriptĐại học Bạc Liêu Truyền thông100% (1)
- Pavement Design1Document57 pagesPavement Design1Mobin AhmadNo ratings yet
- Waterstop TechnologyDocument69 pagesWaterstop TechnologygertjaniNo ratings yet
- Digital MetersDocument47 pagesDigital MetersherovhungNo ratings yet
- Assessment of Students' Oral Communication in English ClassDocument10 pagesAssessment of Students' Oral Communication in English ClassKeebeek S ArbasNo ratings yet
- Carriage RequirementsDocument63 pagesCarriage RequirementsFred GrosfilerNo ratings yet
- Organization and Management Module 3: Quarter 1 - Week 3Document15 pagesOrganization and Management Module 3: Quarter 1 - Week 3juvelyn luegoNo ratings yet
- AlpaGasus: How To Train LLMs With Less Data and More AccuracyDocument6 pagesAlpaGasus: How To Train LLMs With Less Data and More AccuracyMy SocialNo ratings yet
- FDA Approves First Gene Therapy, Betibeglogene Autotemcel (Zynteglo), For Beta-ThalassemiaDocument3 pagesFDA Approves First Gene Therapy, Betibeglogene Autotemcel (Zynteglo), For Beta-ThalassemiaGiorgi PopiashviliNo ratings yet
- Broken BondsDocument20 pagesBroken Bondsapi-316744816No ratings yet
- Product CatalogsDocument12 pagesProduct Catalogscab666No ratings yet
- Spesifikasi PM710Document73 pagesSpesifikasi PM710Phan'iphan'No ratings yet
- E MudhraDownload HardDocument17 pagesE MudhraDownload HardVivek RajanNo ratings yet
- Opc PPT FinalDocument22 pagesOpc PPT FinalnischalaNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (110)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignFrom EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignRating: 4.5 out of 5 stars4.5/5 (39)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (596)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (16)