You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5814)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- SHEM 10 Incident Reporting Rev 00 Jan 07 1Document54 pagesSHEM 10 Incident Reporting Rev 00 Jan 07 1Anonymous iI88LtNo ratings yet
- Portable Power Tools - Risk Assessment2Document3 pagesPortable Power Tools - Risk Assessment2Basit Nawaz100% (1)
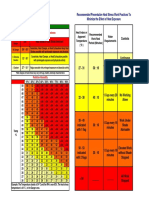
- Heat Stress ChartDocument1 pageHeat Stress ChartJun WangNo ratings yet
- Wahyuningrum Indah Saraswati-01206321A - Tugas P.farprak - Penyimpanan ObatDocument50 pagesWahyuningrum Indah Saraswati-01206321A - Tugas P.farprak - Penyimpanan ObatSaraswati Indah ArumNo ratings yet
- UNIT 4 Review of Related Literature and Studies PDFDocument7 pagesUNIT 4 Review of Related Literature and Studies PDFJohn Lester M. Dela CruzNo ratings yet
- Exposure To Psychosocial Risk Factors in The ConteDocument15 pagesExposure To Psychosocial Risk Factors in The ConteMagally RinconNo ratings yet
- Management of Acquired Defects 2Document50 pagesManagement of Acquired Defects 2Sidhant SudanNo ratings yet
- Slosson Intelligence Test (Sit) Interpretation Examiner: M.A.B Identifying InformationDocument1 pageSlosson Intelligence Test (Sit) Interpretation Examiner: M.A.B Identifying InformationFarrukh Ali KhanNo ratings yet
- Air Quality Index (Aqi) of Lahore Recorded With Aqms On 22.05.2022 (Based On Previous 24 Hours Data)Document1 pageAir Quality Index (Aqi) of Lahore Recorded With Aqms On 22.05.2022 (Based On Previous 24 Hours Data)fahadNo ratings yet
- Satish Kumar Passyavula: E-ISSN: 2663-2268 P-ISSN: 2663-225XDocument2 pagesSatish Kumar Passyavula: E-ISSN: 2663-2268 P-ISSN: 2663-225XMamata BeheraNo ratings yet
- Allison Malm Hs Pe ResumeDocument2 pagesAllison Malm Hs Pe Resumeapi-334423293No ratings yet
- Chapter 5 Statement of The ProblemDocument21 pagesChapter 5 Statement of The ProblemJanzell Anne Borja AlbaniaNo ratings yet
- KOC HSE Sup Action ItemsDocument3 pagesKOC HSE Sup Action ItemsHassan KhattabNo ratings yet
- Soal US XiiDocument9 pagesSoal US Xiimekmekkonkon6No ratings yet
- PC55 JosDocument2 pagesPC55 JosilhamNo ratings yet
- Contoh Tabel ObservasiDocument7 pagesContoh Tabel ObservasinadiyaNo ratings yet
- Jha Lighting ArrestorDocument3 pagesJha Lighting ArrestorRavi thokalNo ratings yet
- Jennifer Jiyoung Hwang Resume 2020-12-09Document3 pagesJennifer Jiyoung Hwang Resume 2020-12-09api-534697316No ratings yet
- 5 ISTFP Conference: International Society of Transference Focused PsychotherapyDocument12 pages5 ISTFP Conference: International Society of Transference Focused PsychotherapyBessy SpNo ratings yet
- Alfeche Kathleen Rose BSN 1 E TFN Case Application Interactive TheoriesDocument4 pagesAlfeche Kathleen Rose BSN 1 E TFN Case Application Interactive TheoriesAiza CeciliaNo ratings yet
- Detox TrainingDocument21 pagesDetox TrainingThinh Do50% (2)
- Mobocertinib (TAK-788) : A Targeted Inhibitor of EGFR Exon 20 Insertion Mutants in Non-Small Cell Lung CancerDocument17 pagesMobocertinib (TAK-788) : A Targeted Inhibitor of EGFR Exon 20 Insertion Mutants in Non-Small Cell Lung CancerKlinikdr RIDHANo ratings yet
- Pollution and Its TypeDocument21 pagesPollution and Its TypeBhavya TiwariNo ratings yet
- Module 3 Drug AwarenessDocument5 pagesModule 3 Drug AwarenessTariga, Dharen Joy J.No ratings yet
- Consenso Rehabilitacion IOIDocument6 pagesConsenso Rehabilitacion IOIMatias Soto ParraNo ratings yet
- Sonia John 2Document1 pageSonia John 2api-534899450No ratings yet
- Preprosthetic SurgeryDocument21 pagesPreprosthetic SurgeryAda MilanovićNo ratings yet
- 2008, Editrice Kurtis: Square-Stepping Exercise Versus Strength and Balance Training For Fall Risk FactorsDocument6 pages2008, Editrice Kurtis: Square-Stepping Exercise Versus Strength and Balance Training For Fall Risk Factorsnandhini raguNo ratings yet
- Benchmark - Ethical Decision MakingDocument6 pagesBenchmark - Ethical Decision Makingapi-520935286No ratings yet
- FLC-L-BM-T2 - Alice JacksonDocument10 pagesFLC-L-BM-T2 - Alice Jacksonvoldemort killerNo ratings yet