You might also like
- Pollination: The Enduring Relationship between Plant and PollinatorFrom EverandPollination: The Enduring Relationship between Plant and PollinatorNo ratings yet
- Papaya SeedDocument6 pagesPapaya SeedAngelineBagtasNo ratings yet
- 483-Article Text-2635-1-10-20201119Document8 pages483-Article Text-2635-1-10-20201119UMT JournalsNo ratings yet
- Leafcutter Ants Adjust Foraging Behaviours When Exposed To Noise DisturbanceDocument11 pagesLeafcutter Ants Adjust Foraging Behaviours When Exposed To Noise DisturbanceTheresa Anne MatunanNo ratings yet
- Agrawal 1999Document8 pagesAgrawal 1999ycamiloguzmansNo ratings yet
- Anaerobic Fungi (Phylum Neocallimastigomycota) : Advances in Understanding Their Taxonomy, Life Cycle, Ecology, Role and Biotechnological PotentialDocument17 pagesAnaerobic Fungi (Phylum Neocallimastigomycota) : Advances in Understanding Their Taxonomy, Life Cycle, Ecology, Role and Biotechnological PotentialAlwi FahroziNo ratings yet
- Jung & Park 2014Document4 pagesJung & Park 2014Carlos Alberto Hector FlechtmannNo ratings yet
- Diversity of Butterflies in Mixed and Pure Plantation AreasDocument4 pagesDiversity of Butterflies in Mixed and Pure Plantation AreasvanpointNo ratings yet
- Competition Among Visitors To Extra Oral NectariesDocument12 pagesCompetition Among Visitors To Extra Oral NectariesjlavilaNo ratings yet
- How To Be An Ant On FigsDocument12 pagesHow To Be An Ant On Figsafira1809No ratings yet
- Pine Trees and BeetlesDocument12 pagesPine Trees and BeetlesSergio D. PérezNo ratings yet
- Ecological and Biotechnological Aspects of LichensDocument12 pagesEcological and Biotechnological Aspects of LichensJosé David QuevedoNo ratings yet
- Botanical Journal of The Linnean Society - 2014 - Ren - Which Food Mimic Floral Traits and Environmental Factors InfluenceDocument13 pagesBotanical Journal of The Linnean Society - 2014 - Ren - Which Food Mimic Floral Traits and Environmental Factors Influence罗依婷No ratings yet
- Hormiga Zombie FosilDocument4 pagesHormiga Zombie FosilUriel RodríguezNo ratings yet
- 121 1995 Biodiversity at Its UtmostDocument14 pages121 1995 Biodiversity at Its UtmostArghya PaulNo ratings yet
- 10.1055@s 2006 924177Document8 pages10.1055@s 2006 924177sayyed bassir ajellehNo ratings yet
- Journal of Applied Ecology - 2022 - Buxton - A Global Review Demonstrating The Importance of Nocturnal Pollinators For CropDocument12 pagesJournal of Applied Ecology - 2022 - Buxton - A Global Review Demonstrating The Importance of Nocturnal Pollinators For CropHenry DzulNo ratings yet
- Gleaning Bats As Underestimated Predators of Herbivorous Insects: Diet of Micronycteris Microtis (Phyllostomidae) in PanamaDocument10 pagesGleaning Bats As Underestimated Predators of Herbivorous Insects: Diet of Micronycteris Microtis (Phyllostomidae) in PanamaTiate TdatdnttNo ratings yet
- Constantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaDocument11 pagesConstantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaindahdwirahNo ratings yet
- Published Paper On Durian Pollination1Document9 pagesPublished Paper On Durian Pollination1Putchong SaraNo ratings yet
- Flores Generalistas, Biodiversidad y Florivoria, Implicaciones para El Origen de Las AngiospermasDocument6 pagesFlores Generalistas, Biodiversidad y Florivoria, Implicaciones para El Origen de Las AngiospermasRobinNo ratings yet
- 10 FAY Conservation of Rare and Endangered Plants Using in Vitro MethodsDocument5 pages10 FAY Conservation of Rare and Endangered Plants Using in Vitro MethodsRebeca GonzagaNo ratings yet
- Community Structure of Sporulating Fungi On Decaying Litters of Shorea SPPDocument8 pagesCommunity Structure of Sporulating Fungi On Decaying Litters of Shorea SPPSitifatimahNo ratings yet
- 10 5923 J Microbiology 20150504 03Document9 pages10 5923 J Microbiology 20150504 03Muhammad Zhafran Iffat Taqy ArifNo ratings yet
- Abdala y Parra Tabla 2005 With Cover Page v2Document8 pagesAbdala y Parra Tabla 2005 With Cover Page v2Zinsou KOUKPONOUNo ratings yet
- TMP 3 D64Document9 pagesTMP 3 D64FrontiersNo ratings yet
- African Parasitoid Fig Wasp Diversification Is A F PDFDocument14 pagesAfrican Parasitoid Fig Wasp Diversification Is A F PDFDaniela lizeth Garzon ClavijoNo ratings yet
- Venus Flytrap Rarely Traps Its PollinatorsDocument9 pagesVenus Flytrap Rarely Traps Its PollinatorsVivi TorresNo ratings yet
- Ecs2 2943Document15 pagesEcs2 2943Aziz Ait AkkaNo ratings yet
- Diversity and Abundance of Lepidopteran Stem Borers and Their Host Plants in EthiopiaDocument13 pagesDiversity and Abundance of Lepidopteran Stem Borers and Their Host Plants in EthiopiaMuluken GoftishuNo ratings yet
- Microbial Diversity: The Importance of Exploration and ConservationDocument6 pagesMicrobial Diversity: The Importance of Exploration and ConservationRedd ZhuangNo ratings yet
- Arnold, Lutzoni - 2007 - Diversity and Host Range of Foliar Fungal Endophytes Are Tropical Leaves Biodiversity HotspotsDocument9 pagesArnold, Lutzoni - 2007 - Diversity and Host Range of Foliar Fungal Endophytes Are Tropical Leaves Biodiversity HotspotsLavinia BarbuNo ratings yet
- Some Biological Features of The Crustacean TypographerDocument13 pagesSome Biological Features of The Crustacean TypographerIJAR JOURNALNo ratings yet
- Butterfly Diversity of NBRI (National Botanical Research Institute), Lucknow, Uttar Pradesh, IndiaDocument7 pagesButterfly Diversity of NBRI (National Botanical Research Institute), Lucknow, Uttar Pradesh, IndiaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Wiley Ecology: This Content Downloaded From 104.239.165.217 On Sun, 26 Jun 2016 18:24:33 UTCDocument10 pagesWiley Ecology: This Content Downloaded From 104.239.165.217 On Sun, 26 Jun 2016 18:24:33 UTCMaría José TorallaNo ratings yet
- J Funeco 2021 101087Document12 pagesJ Funeco 2021 101087Raymundo Lopez NNo ratings yet
- Mokany 2003 OecologiaDocument6 pagesMokany 2003 OecologiaConnie Joy CalawagNo ratings yet
- Tussenbroek Et Al., 2012Document7 pagesTussenbroek Et Al., 2012Bella MorteNo ratings yet
- Amancio Et Al. - 2019 - Feeding Specialization of Flies (Diptera RichardiDocument7 pagesAmancio Et Al. - 2019 - Feeding Specialization of Flies (Diptera Richardigualu_pi_ta_No ratings yet
- Sabu 2010Document18 pagesSabu 2010Sarai Olmedo CruzNo ratings yet
- (2013) Melittophily and OrnithochoryDocument11 pages(2013) Melittophily and OrnithochoryWILLIAM ENRIQUE FIGUEROA MARTINEZNo ratings yet
- Morphological Diversity of Ascobolus and Pilobolus Fungi From Wild Herbivore Dung in Nairobi National Park, KenyaDocument9 pagesMorphological Diversity of Ascobolus and Pilobolus Fungi From Wild Herbivore Dung in Nairobi National Park, KenyaXxxQ.YdhrastNo ratings yet
- Flora: Stavroula Mamoucha, Nikolas Fokialakis, Nikolaos S. ChristodoulakisDocument11 pagesFlora: Stavroula Mamoucha, Nikolas Fokialakis, Nikolaos S. ChristodoulakisRachmat Wihandana AgasiNo ratings yet
- Little 2006Document6 pagesLittle 2006Gigi BarraganNo ratings yet
- DKE215 ch4Document58 pagesDKE215 ch4GUIDO ERNESTO VILLOTA CALVACHINo ratings yet
- 6081 ArticleText 17468 1 10 20121220Document17 pages6081 ArticleText 17468 1 10 20121220Kania TrieswandaNo ratings yet
- Ectoparasites Associated With The Bushveld Gerbil Gerbilliscus Leucogaster and The Role of The Host and Habitat in Shaping Ectoparasite Diversity and InfestationsDocument13 pagesEctoparasites Associated With The Bushveld Gerbil Gerbilliscus Leucogaster and The Role of The Host and Habitat in Shaping Ectoparasite Diversity and InfestationswellingtonlimaNo ratings yet
- 8902 23627 1 PBDocument11 pages8902 23627 1 PBNoorFadilahNo ratings yet
- A Review of Biology and Control of Rhyzopertha DominicaDocument18 pagesA Review of Biology and Control of Rhyzopertha DominicaMasyitah AdianNo ratings yet
- Oomycota GroupDocument7 pagesOomycota GroupZaman KhanNo ratings yet
- JM Cocos,+Jurnal+Tegar+ProkDocument8 pagesJM Cocos,+Jurnal+Tegar+ProkbuanaNo ratings yet
- Diversity and Ef Ciency of Wild Pollinators of Watermelon (Citrullus Lanatus (Thunb.) Mansf.) at Yatta (Kenya)Document7 pagesDiversity and Ef Ciency of Wild Pollinators of Watermelon (Citrullus Lanatus (Thunb.) Mansf.) at Yatta (Kenya)Shailendra RajanNo ratings yet
- Boone 2008Document13 pagesBoone 2008joaopauloramosdemeloNo ratings yet
- Palynology of Some Species in The Family Cucurbitaceae From NigeriaDocument11 pagesPalynology of Some Species in The Family Cucurbitaceae From NigeriaIJAR JOURNALNo ratings yet
- Bioaccumulation of The Artificial Cs-137 and The Natural Radionuclides Th-234, Ra-226, K-40 in The Fruit Bodies of Basidiomycetes in Greece - 2015Document13 pagesBioaccumulation of The Artificial Cs-137 and The Natural Radionuclides Th-234, Ra-226, K-40 in The Fruit Bodies of Basidiomycetes in Greece - 2015ATBX&MT PhòngNo ratings yet
- Host Specificity in Bat Ectoparasites: A Natural Experiment: ArticleDocument9 pagesHost Specificity in Bat Ectoparasites: A Natural Experiment: Articlefaradisa35No ratings yet
- Diversidad de Hongos Que Destruyen Nematodos en Taita Taveta, KeniaDocument6 pagesDiversidad de Hongos Que Destruyen Nematodos en Taita Taveta, KeniaPaola LopezNo ratings yet
- Research 8Document5 pagesResearch 8Jamil SalibioNo ratings yet
- Codella - 1996 - Individual and Social Components of Wood Ant Response To Conifer SawflyDocument11 pagesCodella - 1996 - Individual and Social Components of Wood Ant Response To Conifer Sawfly永政澤No ratings yet
- Evaluation of Fipronil and Imidacloprid As Bait Active Ingredients Against Fungus-Growing Termites (Blattodea - Termitidae - Macrotermitinae)Document10 pagesEvaluation of Fipronil and Imidacloprid As Bait Active Ingredients Against Fungus-Growing Termites (Blattodea - Termitidae - Macrotermitinae)xidegaNo ratings yet
- Comparative Study of The Antioxidant and Enzyme inDocument21 pagesComparative Study of The Antioxidant and Enzyme inRifqi Fathul ArroisiNo ratings yet
- Reppert and Roode 2018 - HighlightDocument14 pagesReppert and Roode 2018 - HighlightRifqi Fathul ArroisiNo ratings yet
- Introduction To Fish Oil Oxidation, Oxidation Prevention, and Oxidation CorrectionDocument6 pagesIntroduction To Fish Oil Oxidation, Oxidation Prevention, and Oxidation CorrectionRifqi Fathul ArroisiNo ratings yet
- Characterization of Cognitive Deficits in Rats Overexpressing Human Alpha-Synuclein in The Ventral Tegmental Area and Medial Septum Using Recombinant Adeno-Associated Viral Vectors (2013)Document12 pagesCharacterization of Cognitive Deficits in Rats Overexpressing Human Alpha-Synuclein in The Ventral Tegmental Area and Medial Septum Using Recombinant Adeno-Associated Viral Vectors (2013)Rifqi Fathul ArroisiNo ratings yet
- s40071 019 0221 1Document13 pagess40071 019 0221 1Rifqi Fathul ArroisiNo ratings yet
- Animal Behaviour: Alice Bouchard, Klaus ZuberbühlerDocument15 pagesAnimal Behaviour: Alice Bouchard, Klaus ZuberbühlerRifqi Fathul ArroisiNo ratings yet
- Extraction and Microencapsulation of Tuna Virgin FDocument11 pagesExtraction and Microencapsulation of Tuna Virgin FRifqi Fathul ArroisiNo ratings yet
- Molecular Pathogenesis of Shigella SPP.: Controlling Host Cell Signaling, Invasion, and Death by Type III SecretionDocument23 pagesMolecular Pathogenesis of Shigella SPP.: Controlling Host Cell Signaling, Invasion, and Death by Type III SecretionRifqi Fathul ArroisiNo ratings yet
- Microsatellite Loci For The Weaver Ant Oecophylla SmaragdinaDocument4 pagesMicrosatellite Loci For The Weaver Ant Oecophylla SmaragdinaRifqi Fathul ArroisiNo ratings yet
- Soil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemDocument25 pagesSoil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemRifqi Fathul ArroisiNo ratings yet
- Healthcare 09 01536Document11 pagesHealthcare 09 01536Rifqi Fathul ArroisiNo ratings yet
- Shigella Pathogenesis: Molecular and Computational Insights: ReviewDocument23 pagesShigella Pathogenesis: Molecular and Computational Insights: ReviewRifqi Fathul ArroisiNo ratings yet
- Genome-Scale Metabolic Modeling Reveals Sars-Cov-2-Induced Metabolic Changes and Antiviral TargetsDocument19 pagesGenome-Scale Metabolic Modeling Reveals Sars-Cov-2-Induced Metabolic Changes and Antiviral TargetsRifqi Fathul ArroisiNo ratings yet
- Teow Et Al (2016) - Curcumin As AntibacterialDocument10 pagesTeow Et Al (2016) - Curcumin As AntibacterialYuyun Aryuni MusrifNo ratings yet
- Crispr Cas9 Nobel Chemistry November 2020Document34 pagesCrispr Cas9 Nobel Chemistry November 2020Rifqi Fathul ArroisiNo ratings yet
- Grismer Et Al 2016 Ansonia KhaochangensisDocument26 pagesGrismer Et Al 2016 Ansonia KhaochangensisRifqi Fathul ArroisiNo ratings yet
- Comparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Document24 pagesComparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Rifqi Fathul ArroisiNo ratings yet
- Species Diversity, Elevational Distribution and Reproductive Modes in An Amphibian Community at The Matang Range, Sarawak (Borneo)Document35 pagesSpecies Diversity, Elevational Distribution and Reproductive Modes in An Amphibian Community at The Matang Range, Sarawak (Borneo)Rifqi Fathul ArroisiNo ratings yet
- SubsDocument15 pagesSubsRifqi Fathul ArroisiNo ratings yet
- Ansonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupDocument9 pagesAnsonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupRifqi Fathul ArroisiNo ratings yet
- Ansonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupDocument9 pagesAnsonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupRifqi Fathul ArroisiNo ratings yet
- Comparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Document24 pagesComparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Rifqi Fathul ArroisiNo ratings yet
- Cosmetics 08 00004 v2Document13 pagesCosmetics 08 00004 v2Rifqi Fathul ArroisiNo ratings yet
- Aquatic Ecosystem and Biodiversity A ReviewDocument13 pagesAquatic Ecosystem and Biodiversity A ReviewRifqi Fathul ArroisiNo ratings yet
- Characterization of The Chemical Composition of Adenostemma Lavenia (L.) Kuntze and Adenostemma Platyphyllum CassDocument9 pagesCharacterization of The Chemical Composition of Adenostemma Lavenia (L.) Kuntze and Adenostemma Platyphyllum Casszan_anwarNo ratings yet
- Foods 09 00073Document12 pagesFoods 09 00073Rifqi Fathul ArroisiNo ratings yet
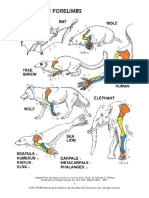
- Adapted From The Human Evolution Coloring Book, 2D Ed., by Adrienne L. Zihlman. Produced by Coloring Concepts Inc. New York: Harpercollins, 2001Document1 pageAdapted From The Human Evolution Coloring Book, 2D Ed., by Adrienne L. Zihlman. Produced by Coloring Concepts Inc. New York: Harpercollins, 2001Rifqi Fathul ArroisiNo ratings yet
- Color1 - Molecular CalibrationDocument1 pageColor1 - Molecular CalibrationRifqi Fathul ArroisiNo ratings yet
- References: Milda Yulia Solihah, 2016Document7 pagesReferences: Milda Yulia Solihah, 2016Rifqi Fathul ArroisiNo ratings yet
- Catalyst 3750 Series Switches TroubleshootDocument19 pagesCatalyst 3750 Series Switches TroubleshootSugumar DuraisamyNo ratings yet
- Uttar Pradesh Universities Act 1973Document73 pagesUttar Pradesh Universities Act 1973ifjosofNo ratings yet
- Bootstrap Aggregating Multivariate Adaptive Regression Spline For Observational Studies in Diabetes CasesDocument8 pagesBootstrap Aggregating Multivariate Adaptive Regression Spline For Observational Studies in Diabetes CasesTika MijayantiNo ratings yet
- Nursing Documentation Course 2020Document36 pagesNursing Documentation Course 2020Marianne Laylo100% (1)
- 11v.jigisha Chaptear2Document53 pages11v.jigisha Chaptear2Anirban PalNo ratings yet
- Statistics On Lgbtiq Inclusion in NorwayDocument46 pagesStatistics On Lgbtiq Inclusion in NorwayНиколай ВавиличевNo ratings yet
- 1 2 4 Ak Sequentiallogicdesign Counters DLBDocument7 pages1 2 4 Ak Sequentiallogicdesign Counters DLBapi-290804719100% (1)
- Impulsive Buying PDFDocument146 pagesImpulsive Buying PDFrukwavuNo ratings yet
- Operations Research Letters: Meichun Lin, Woonghee Tim Huh, Guohua WanDocument8 pagesOperations Research Letters: Meichun Lin, Woonghee Tim Huh, Guohua WanQuỳnh NguyễnNo ratings yet
- IC HDL Lab ManualDocument82 pagesIC HDL Lab ManualRakshitha AngelNo ratings yet
- Types of Non Verbal CommunicationDocument7 pagesTypes of Non Verbal Communicationgaurav gharat100% (1)
- Spelling Menu Days and MonthsDocument1 pageSpelling Menu Days and MonthsLisl WindhamNo ratings yet
- Ponty Maurice (1942,1968) Structure of BehaviorDocument131 pagesPonty Maurice (1942,1968) Structure of BehaviorSnorkel7No ratings yet
- Unit 1session 2 Performance MGMTDocument38 pagesUnit 1session 2 Performance MGMTMayank TrivediNo ratings yet
- Khutbah About The QuranDocument3 pagesKhutbah About The QurantakwaniaNo ratings yet
- 1386258018727Document21 pages1386258018727Roberto MuñozNo ratings yet
- Management of Liver Trauma in Adults: Nasim Ahmed, Jerome J VernickDocument7 pagesManagement of Liver Trauma in Adults: Nasim Ahmed, Jerome J VernickwiraNo ratings yet
- Seerat Mujaddid Alf-e-Sani (Urdu)Document518 pagesSeerat Mujaddid Alf-e-Sani (Urdu)Talib Ghaffari100% (12)
- 07.03.09 Chest PhysiotherapyDocument10 pages07.03.09 Chest PhysiotherapyMuhammad Fuad MahfudNo ratings yet
- Comprehensive Compressed Air Assessments: The 5-Step ProcessDocument8 pagesComprehensive Compressed Air Assessments: The 5-Step ProcessANDRESNo ratings yet
- Low Intermediate Korean Vocabulary and GrammarDocument10 pagesLow Intermediate Korean Vocabulary and GrammarTuong Van Nguyen100% (3)
- BTCTL 17Document5 pagesBTCTL 17Alvin BenaventeNo ratings yet
- 5568 AssignmentDocument12 pages5568 AssignmentAtif AliNo ratings yet
- Universal Declaration of Human Rights - United NationsDocument12 pagesUniversal Declaration of Human Rights - United NationsSafdar HussainNo ratings yet
- Physical Education 10 WEEK 2Document10 pagesPhysical Education 10 WEEK 2Israel MarquezNo ratings yet
- HF CharactersDocument5 pagesHF CharactersAudri DebnathNo ratings yet
- Shalini NaagarDocument2 pagesShalini NaagarAazam AdtechiesNo ratings yet
- Sales Purchases Returns Day BookDocument8 pagesSales Purchases Returns Day BookAung Zaw HtweNo ratings yet
- Selfishness EssayDocument8 pagesSelfishness Essayuiconvbaf100% (2)
- مذكرة التأسيس الرائعة لغة انجليزية للمبتدئين?Document21 pagesمذكرة التأسيس الرائعة لغة انجليزية للمبتدئين?Manar SwaidanNo ratings yet