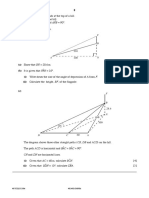
You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5807)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (346)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Pre AlgebraDocument706 pagesPre AlgebraMLSBU11100% (1)
- Coffee Marketing PlanDocument24 pagesCoffee Marketing PlanAnkit Goswami100% (1)
- General Psychology by Amit AbrahamDocument596 pagesGeneral Psychology by Amit AbrahamAnni Zomiyyah S1 Ilmu PerpustakaanNo ratings yet
- Jyotish - Ersnt Wilhem - Upadesa Sutras 1Document122 pagesJyotish - Ersnt Wilhem - Upadesa Sutras 1pm plassanalNo ratings yet
- GB2 Q2 W7Document17 pagesGB2 Q2 W7qhuenzielNo ratings yet
- Abruptio Placenta NCP 2 FinalDocument19 pagesAbruptio Placenta NCP 2 FinalTin100% (1)
- Ficha Lobisomem O Apocalipse Editavel PDFDocument4 pagesFicha Lobisomem O Apocalipse Editavel PDFRadazz Kiol100% (1)
- A Qualitative Study About Mood-Boosting of Senior Highschool in Our Lady of Fatima UniversityDocument7 pagesA Qualitative Study About Mood-Boosting of Senior Highschool in Our Lady of Fatima UniversityPrince Ariean Songcuan PinedaNo ratings yet
- Betamate 2090Document4 pagesBetamate 2090José Antonio CuberoNo ratings yet
- BÀI TẬP CODE GOOGLEDocument2 pagesBÀI TẬP CODE GOOGLETrần Phúc QuýNo ratings yet
- I. Objectives: Fort Pilar ST., Zamboanga CityDocument10 pagesI. Objectives: Fort Pilar ST., Zamboanga CityDanizelle Kaye Cadocoy BernardoNo ratings yet
- Ema Competency Framework - enDocument67 pagesEma Competency Framework - enfungcheu7193No ratings yet
- Orthopedo AssDocument4 pagesOrthopedo AssIvyRoselleLacasandileCabañeroNo ratings yet
- Murdoch University Auslandssemester Study Abroad BrochureDocument30 pagesMurdoch University Auslandssemester Study Abroad BrochureEdson PrataNo ratings yet
- The Ancient Andean World 3.982 Fall 2018 Syllabus: Lechtman@mit - EduDocument5 pagesThe Ancient Andean World 3.982 Fall 2018 Syllabus: Lechtman@mit - EduShubhangaNo ratings yet
- TRIGONOMETRY 99 To 2013 All Questions PDFDocument87 pagesTRIGONOMETRY 99 To 2013 All Questions PDFfalak fatima100% (1)
- Organization Structure/Enterprises Hierarchy: Diagram:01Document4 pagesOrganization Structure/Enterprises Hierarchy: Diagram:01Pranitkumar pathadeNo ratings yet
- Report 1535390498197152524Document14 pagesReport 1535390498197152524Geca importaciones Geca ImportacionesNo ratings yet
- 7 - Milberg e Winkler - Outsourcing EconomicsDocument378 pages7 - Milberg e Winkler - Outsourcing EconomicsDaniel CarvalhoNo ratings yet
- Deep Tubewell 076.77Document28 pagesDeep Tubewell 076.77Civil EngineeringNo ratings yet
- Ina 114Document17 pagesIna 114Moditha LakshanNo ratings yet
- Welfare Facilities 4Document5 pagesWelfare Facilities 4Manoj ManojNo ratings yet
- Challenges and Scope of Data Science ProjectDocument21 pagesChallenges and Scope of Data Science ProjectMihir PesswaniNo ratings yet
- Method Statement: Site Set Up: Compound, Parking & Material StorageDocument19 pagesMethod Statement: Site Set Up: Compound, Parking & Material StorageVICTORNo ratings yet
- 1 Leaner - S Style EQUIBALDocument3 pages1 Leaner - S Style EQUIBALDaniel Florence EquibalNo ratings yet
- Gov. Pablo Borbon Campus II, Alangilan, Batangas City, Philippines 4200 WWW - Batstate-U.edu - PH Telefax: (043) 300-4044 Locs. 106-108Document6 pagesGov. Pablo Borbon Campus II, Alangilan, Batangas City, Philippines 4200 WWW - Batstate-U.edu - PH Telefax: (043) 300-4044 Locs. 106-108Olibear ArellanoNo ratings yet
- 11521i PDFDocument1 page11521i PDFNisa Javadd0% (1)
- SM-J400F Common Tshoo 7Document45 pagesSM-J400F Common Tshoo 7Cristian Dalla RivaNo ratings yet
- Cap 10 Interpretation of Granitc RocksDocument13 pagesCap 10 Interpretation of Granitc Rocks6m9jrdfyv9No ratings yet
- Final Assessment For Laboratoires de Biarritz Marault Coudrin Colaianni Capelle Chatelain VlajcicDocument27 pagesFinal Assessment For Laboratoires de Biarritz Marault Coudrin Colaianni Capelle Chatelain VlajcicLeaaa MNo ratings yet