You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Grade 6Document18 pagesGrade 6Cht GebyNo ratings yet
- Cat Oxsilan InternetDocument49 pagesCat Oxsilan InternetAnonymous 1vuTwJVDjYNo ratings yet
- Revealed Comparative AdvantageDocument5 pagesRevealed Comparative AdvantageiamrahulkumarNo ratings yet
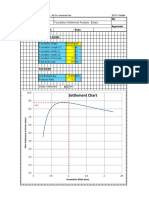
- Settlement Chart: Foundation Settlement Analysis - ElasticDocument2 pagesSettlement Chart: Foundation Settlement Analysis - ElasticCSEC Uganda Ltd.No ratings yet
- Lab Guide: Musculoskeletal AssessmentDocument6 pagesLab Guide: Musculoskeletal AssessmentDaisy MellaNo ratings yet
- 4aa4 1063enwDocument8 pages4aa4 1063enwHenderson and SantosNo ratings yet
- Easy Pinjaman Ekspres PDSDocument4 pagesEasy Pinjaman Ekspres PDSFizz FirdausNo ratings yet
- Import FinalDocument5 pagesImport FinalRaza AliNo ratings yet
- B - 21 Raider (6th Gen Fighter)Document2 pagesB - 21 Raider (6th Gen Fighter)AbhijeetNo ratings yet
- DS - 20190709 - E2 - E2 198S-264S Datasheet - V10 - ENDocument13 pagesDS - 20190709 - E2 - E2 198S-264S Datasheet - V10 - ENCristina CorfaNo ratings yet
- Doerrenberg Steel Foundry EnglishDocument16 pagesDoerrenberg Steel Foundry EnglishAditya PratapNo ratings yet
- Milk Production Trend - Dairy Development - Outlook PDFDocument10 pagesMilk Production Trend - Dairy Development - Outlook PDFrubel_nsuNo ratings yet
- Delta Background-Group AssignmentDocument2 pagesDelta Background-Group AssignmentTinashe100% (2)
- Reference Data Types in JavaDocument3 pagesReference Data Types in Javaeyn batallaNo ratings yet
- AMPLA Twilight Seminar JORC PresentationDocument65 pagesAMPLA Twilight Seminar JORC Presentationarief_7No ratings yet
- Catalogue of Jumbo: Jumbo Power Technology Co.,LimitedDocument20 pagesCatalogue of Jumbo: Jumbo Power Technology Co.,LimitedJosé Daniel Interventoria AP MocoaNo ratings yet
- Mathematical LanguageDocument9 pagesMathematical Languagejrqagua00332No ratings yet
- HSE Manager JDDocument2 pagesHSE Manager JDRukun SinagaNo ratings yet
- MIL-H-6875H Heat Treatment of SteelDocument29 pagesMIL-H-6875H Heat Treatment of Steelgoathead61No ratings yet
- Module 4 - Design of Tension MembersDocument30 pagesModule 4 - Design of Tension MembersSreelakshmi GNo ratings yet
- Union Feb. 10, 2016Document16 pagesUnion Feb. 10, 2016Your News. When You Want It.No ratings yet
- Nodeep Kaur The Jailed Activist Meena Harris Tweeted AboutDocument11 pagesNodeep Kaur The Jailed Activist Meena Harris Tweeted AboutBIDIFianNo ratings yet
- OTPBank Securities SI 20210217Document6 pagesOTPBank Securities SI 20210217bdsourav0No ratings yet
- 12th Commerce Study Material English MediumDocument97 pages12th Commerce Study Material English MediumDeepak ajar100% (1)
- Exam Preparation Chartered Member Solutions 20080403Document36 pagesExam Preparation Chartered Member Solutions 20080403Jordy NgNo ratings yet
- Production Technology of Cow PeaDocument20 pagesProduction Technology of Cow PeaMUHAMMAD AQEELNo ratings yet
- 01 AR PFMEA - TemplateDocument3 pages01 AR PFMEA - TemplateAndrew DoNo ratings yet
- Mapeh-Arts: Quarter 2 - Module 2: Technology-Based ArtDocument24 pagesMapeh-Arts: Quarter 2 - Module 2: Technology-Based ArtTrecia Roxas SajolNo ratings yet
- Annexure 5.14 BC Mix DesignDocument10 pagesAnnexure 5.14 BC Mix Designmiesty50% (2)
- Kagan Quiz Quiz TradeDocument2 pagesKagan Quiz Quiz Tradeapi-401932164No ratings yet