You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Book (Prayer)Document232 pagesBook (Prayer)mintillaNo ratings yet
- Chapter 1 Bio320 PDFDocument3 pagesChapter 1 Bio320 PDFNurul Amirah HossainNo ratings yet
- A Systematic Review On The Comparison of Molecular Gene Editing ToolsDocument8 pagesA Systematic Review On The Comparison of Molecular Gene Editing ToolsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Future Libraries DigitalDocument44 pagesFuture Libraries DigitalmintillaNo ratings yet
- Entropy 16 05428 v2Document95 pagesEntropy 16 05428 v2mintillaNo ratings yet
- Djordjevic 2007Document7 pagesDjordjevic 2007mintillaNo ratings yet
- Rao 2011Document10 pagesRao 2011mintillaNo ratings yet
- Fractal Space-Time and Microphysics Towards A Theory of - Nottale, Laurent - 1993Document345 pagesFractal Space-Time and Microphysics Towards A Theory of - Nottale, Laurent - 1993mintillaNo ratings yet
- Aspects of Topology - Christenson, Charles O Voxman, William L - 1998Document518 pagesAspects of Topology - Christenson, Charles O Voxman, William L - 1998mintillaNo ratings yet
- ButterfieldDocument148 pagesButterfieldmintillaNo ratings yet
- Peskin1982 PDFDocument2 pagesPeskin1982 PDFmintillaNo ratings yet
- Upon Opening The Black Box and Finding It Empty SoDocument19 pagesUpon Opening The Black Box and Finding It Empty SomintillaNo ratings yet
- Yajima 1985Document7 pagesYajima 1985mintillaNo ratings yet
- Beaulieu 2006Document5 pagesBeaulieu 2006mintillaNo ratings yet
- Cancer ChemotherapyDocument1 pageCancer ChemotherapymintillaNo ratings yet
- Handbook of Cryo-Preparation Methods For Electron Microscopy (Methods in Visualization) (Annie Cavalier, Daniele Spehner Etc.Document630 pagesHandbook of Cryo-Preparation Methods For Electron Microscopy (Methods in Visualization) (Annie Cavalier, Daniele Spehner Etc.mintillaNo ratings yet
- Barnes and Bloor Rationality and RelativismDocument16 pagesBarnes and Bloor Rationality and RelativismmintillaNo ratings yet
- Listening Comprehension IDocument14 pagesListening Comprehension ImintillaNo ratings yet
- Kinetic Investigation of Eu (III) - Humate Interactions by Ion Exchange ResinsDocument7 pagesKinetic Investigation of Eu (III) - Humate Interactions by Ion Exchange ResinsmintillaNo ratings yet
- Participate Soil Organic-Matter Changes Across A Grassland Cultivation SequenceDocument7 pagesParticipate Soil Organic-Matter Changes Across A Grassland Cultivation SequencemintillaNo ratings yet
- Zechmeister Boltenstern2015Document23 pagesZechmeister Boltenstern2015mintillaNo ratings yet
- A Re-Investigation of Resveratrol Synthesis by Perkins ReactionDocument7 pagesA Re-Investigation of Resveratrol Synthesis by Perkins ReactionmintillaNo ratings yet
- Journal of Chemical Research Index Jan-Feb2021Document12 pagesJournal of Chemical Research Index Jan-Feb2021mintillaNo ratings yet
- Bindur CheleDocument74 pagesBindur ChelemintillaNo ratings yet
- A Modern Formal Logic Primer - Predicate Theory. (Paul Teller) PDFDocument279 pagesA Modern Formal Logic Primer - Predicate Theory. (Paul Teller) PDFmintillaNo ratings yet
- AP Bio Chapter 12&13 Study GuideDocument6 pagesAP Bio Chapter 12&13 Study Guideben_sorkinNo ratings yet
- Optimal Capacitors Placement in Distribution Networks Using Genetic Algorithm: A Dimension Reducing ApproachDocument7 pagesOptimal Capacitors Placement in Distribution Networks Using Genetic Algorithm: A Dimension Reducing ApproachDavid Omar Torres GutierrezNo ratings yet
- 5 Points 10 Points 15 Points 20 Points: TotalDocument5 pages5 Points 10 Points 15 Points 20 Points: TotalMary Grace AgueteNo ratings yet
- WES WGS Brochure Pages V2.1eng Web 20201019Document16 pagesWES WGS Brochure Pages V2.1eng Web 20201019drumerNo ratings yet
- COPDDocument50 pagesCOPDalilnaNo ratings yet
- Week 3: Cell Modifications: General Biology 1Document3 pagesWeek 3: Cell Modifications: General Biology 1Florene Bhon GumapacNo ratings yet
- Fibrous Proteins: BY Dr. Sehrish LodhiDocument31 pagesFibrous Proteins: BY Dr. Sehrish Lodhisash34No ratings yet
- Introduction To Cellular AberrationDocument11 pagesIntroduction To Cellular AberrationAdiel CalsaNo ratings yet
- Breeding Guppy - Guppy Guru DerekDocument5 pagesBreeding Guppy - Guppy Guru DerekAnthony JohanNo ratings yet
- Solleza PT2 MatipidDocument3 pagesSolleza PT2 Matipidelah jhaneNo ratings yet
- Fundamental Molecular Biology: GenomesDocument45 pagesFundamental Molecular Biology: GenomesMoonHoLeeNo ratings yet
- Chemical & Life Sciences: School ofDocument40 pagesChemical & Life Sciences: School ofweiweiahNo ratings yet
- SorghumDocument6 pagesSorghumASHUTOSH MOHANTYNo ratings yet
- B04 Joenathan Gabriel Chua - DNA Extraction - Activity PaperDocument7 pagesB04 Joenathan Gabriel Chua - DNA Extraction - Activity PaperB04 Joenathan Gabriel ChuaNo ratings yet
- Biology NotesDocument9 pagesBiology NotesPrasad KhotNo ratings yet
- 2020MB6953RDocument1 page2020MB6953RFaizan RasoolNo ratings yet
- Hiphedera ALDocument154 pagesHiphedera ALMichelle DulayNo ratings yet
- Ovarian ReserverDocument35 pagesOvarian Reserverpramod singhNo ratings yet
- Assignment 03 Henry PriceDocument2 pagesAssignment 03 Henry PriceHenry PriceNo ratings yet
- Immunoserology: Medical Technology Assessment Progam 1 Discussed By: Mr. Mark MendrosDocument8 pagesImmunoserology: Medical Technology Assessment Progam 1 Discussed By: Mr. Mark MendrosLea Juan100% (1)
- HyaluronicAcid May 2021Document23 pagesHyaluronicAcid May 2021a290733No ratings yet
- PHD Studentship - Development of A 3D Printed Biohybrid Lung at Loughborough UniversityDocument3 pagesPHD Studentship - Development of A 3D Printed Biohybrid Lung at Loughborough UniversityOzden IsbilirNo ratings yet
- ES 01 Insightsonindia - com-SYNOPSIS Insights 70 Days Ethics Plan Day 1Document5 pagesES 01 Insightsonindia - com-SYNOPSIS Insights 70 Days Ethics Plan Day 1sunilNo ratings yet
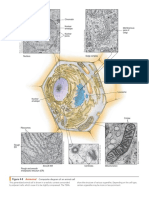
- Figure 4-8 Composite Diagram of An Animal Cell: AnimatedDocument1 pageFigure 4-8 Composite Diagram of An Animal Cell: AnimatedElrey AnatolNo ratings yet
- Biopolymers in Wastewater TreatmentDocument4 pagesBiopolymers in Wastewater Treatmentبارق التجاريةNo ratings yet
- Namma Kalvi 11th Bio-Botany Study Material English Medium 219431Document51 pagesNamma Kalvi 11th Bio-Botany Study Material English Medium 219431Prasanth PrasanthNo ratings yet
- De Guzman OE - 4Document3 pagesDe Guzman OE - 4Jericson De GuzmanNo ratings yet
- TTD LecturersDocument101 pagesTTD LecturersRajesh AppariNo ratings yet