You might also like
- Request For Proposal Construction & Phase 1 OperationDocument116 pagesRequest For Proposal Construction & Phase 1 Operationsobhi100% (2)
- Travis Walton Part 1 MUFON Case FileDocument346 pagesTravis Walton Part 1 MUFON Case FileClaudio Silva100% (1)
- The Medicinal Fungus Cordyceps Militaris: Research and DevelopmentDocument17 pagesThe Medicinal Fungus Cordyceps Militaris: Research and DevelopmentChongtham Allaylay Devi (SBSR Assistant Professor)No ratings yet
- Fyp ProposalDocument5 pagesFyp ProposalEwant RamlanNo ratings yet
- LARVICIDAL EFFECT OF AMPALAYA (Momordica Charantia) FRUIT JUICE ON Aedes MOSQUITO LARVAEDocument53 pagesLARVICIDAL EFFECT OF AMPALAYA (Momordica Charantia) FRUIT JUICE ON Aedes MOSQUITO LARVAERajesh Kumar Asunala57% (7)
- Copy Resit APLC MiniAssignmentDocument5 pagesCopy Resit APLC MiniAssignmentChong yaoNo ratings yet
- Morphology and Phylogeny of Neoscytalidium Sp. Nov. (Botryosphaeriaceae)Document7 pagesMorphology and Phylogeny of Neoscytalidium Sp. Nov. (Botryosphaeriaceae)Alexandre MatiasNo ratings yet
- Debregeasia Mutation HotspotDocument14 pagesDebregeasia Mutation HotspotamitNo ratings yet
- Periodic Acid Schiff Reactions and General Tissue Morphology of Conventionally Processed Versus Two Rapid Microwave Processed TissuesDocument15 pagesPeriodic Acid Schiff Reactions and General Tissue Morphology of Conventionally Processed Versus Two Rapid Microwave Processed TissuesRestiyaniNo ratings yet
- Are Some Endophytes of Musa Acuminata Latent PathoDocument11 pagesAre Some Endophytes of Musa Acuminata Latent PathoJosé Antonio .Thomas ArgüellesNo ratings yet
- Mycological Progress 24 May 2012Document17 pagesMycological Progress 24 May 2012Harendra ChunduruNo ratings yet
- Cordyceps Pruinosa Produces Cordycepin and N6 - (2-Hydroxyethyl) - Adenosine in CultureDocument13 pagesCordyceps Pruinosa Produces Cordycepin and N6 - (2-Hydroxyethyl) - Adenosine in CultureAnh NguyenNo ratings yet
- Labeda Et Al. 2012Document33 pagesLabeda Et Al. 2012Jose FernandoNo ratings yet
- Sajeewa FungalDiversity 2012 1Document36 pagesSajeewa FungalDiversity 2012 1Claudia TolpNo ratings yet
- 10 11648 J Fem 20150102 12Document6 pages10 11648 J Fem 20150102 12kingwinyashNo ratings yet
- Norphanphounetal 2021Document23 pagesNorphanphounetal 2021disha aroraNo ratings yet
- Cryptic diversity in the Pseudo-nitzschia delicatissima complexDocument19 pagesCryptic diversity in the Pseudo-nitzschia delicatissima complexdavidNo ratings yet
- Cordyceps Takaomontana AT LANGBIAN MOUNTAIN,: Discovery of Entomopathogenic Fungi Lam Dong, Viet NamDocument8 pagesCordyceps Takaomontana AT LANGBIAN MOUNTAIN,: Discovery of Entomopathogenic Fungi Lam Dong, Viet NamAnh NguyenNo ratings yet
- Valsaria and The Valsariales: Fungal Diversity May 2015Document45 pagesValsaria and The Valsariales: Fungal Diversity May 2015wilhelmNo ratings yet
- Soisooketal ActaChiro 2015Document17 pagesSoisooketal ActaChiro 2015Dhony ArdiansyahNo ratings yet
- Ayuningrum 2017 Iop Conf Ser 3 A Earth Environ Sci 55 012029 CompressedDocument10 pagesAyuningrum 2017 Iop Conf Ser 3 A Earth Environ Sci 55 012029 Compressedardhanu asagabaldanNo ratings yet
- 2010ActaOceanol SinDocument9 pages2010ActaOceanol SinRifqi Fathul ArroisiNo ratings yet
- CylindrocladiellaDocument35 pagesCylindrocladiellaestefaniacb1305No ratings yet
- 2016vol 73phuping714Document9 pages2016vol 73phuping714Libertine Agatha DensingNo ratings yet
- Advances in The Phylogenesis of Agaricales and ItsDocument9 pagesAdvances in The Phylogenesis of Agaricales and ItsYoyo CastilloNo ratings yet
- AnfíbiosDocument11 pagesAnfíbiosRita PinheiroNo ratings yet
- Preliminary Study On Identification of Spiders Using Mitochondrial DnaDocument3 pagesPreliminary Study On Identification of Spiders Using Mitochondrial DnakashmiraNo ratings yet
- Epidermal_and_Cytological_Studies_on_CulDocument7 pagesEpidermal_and_Cytological_Studies_on_CuladeliasiregarNo ratings yet
- The Species Identity of The Widely Cultivated Ganoderma, G. Lucidum' (Ling-Zhi), in ChinaDocument13 pagesThe Species Identity of The Widely Cultivated Ganoderma, G. Lucidum' (Ling-Zhi), in ChinaTeodoro Chivatá BedoyaNo ratings yet
- Foldscope study of Drosophila reproductive organs and human cellsDocument9 pagesFoldscope study of Drosophila reproductive organs and human cellsJosé Antonio .Thomas ArgüellesNo ratings yet
- The Origin and Evolution of PhototropinsDocument11 pagesThe Origin and Evolution of Phototropinsutama 3002No ratings yet
- PDFDocument13 pagesPDFAricatul JannahNo ratings yet
- EXTRACTION OF DNA FROM CHEEK CELLSDocument8 pagesEXTRACTION OF DNA FROM CHEEK CELLSFiel Cris LansangNo ratings yet
- Arr 2009 Final With LinksDocument63 pagesArr 2009 Final With LinksStarLink1No ratings yet
- Full TextDocument295 pagesFull TextkanNo ratings yet
- Downloadfile PDFDocument12 pagesDownloadfile PDFAhmed AbedoNo ratings yet
- Neurogenetics Lab Record Nitisha SinghDocument45 pagesNeurogenetics Lab Record Nitisha Singhnitisha singhNo ratings yet
- Fungicolous Fungi Terminology, Diversity, Distribution, EvolutionDocument94 pagesFungicolous Fungi Terminology, Diversity, Distribution, Evolutionela.sofiaNo ratings yet
- Zebrafish As A Genetic Model Organism: Biotecnologia Aplicada October 2006Document7 pagesZebrafish As A Genetic Model Organism: Biotecnologia Aplicada October 2006shaheena ahmedNo ratings yet
- In Vitro-Derived Plants of Dendrobium NobileDocument5 pagesIn Vitro-Derived Plants of Dendrobium NobilejammesNo ratings yet
- 795 Flavoparmelia PradoDocument13 pages795 Flavoparmelia PradoSergio Andres Galan AyalaNo ratings yet
- 1035 10VPDocument13 pages1035 10VPmateus silvaNo ratings yet
- Misof_2014Document7 pagesMisof_2014Tainara SobrozaNo ratings yet
- Phylogenetic Classification of Cordyceps and The Clavicipitaceous Fungi. Stud MycolDocument56 pagesPhylogenetic Classification of Cordyceps and The Clavicipitaceous Fungi. Stud MycolNgocGiauNguyenNo ratings yet
- Leonelli2013 - What Is A Model OrganismDocument4 pagesLeonelli2013 - What Is A Model OrganismIonascu AdrianNo ratings yet
- # 1566 Revision Bangiales JP 2011Document34 pages# 1566 Revision Bangiales JP 2011Gabriela Reboreda NavaNo ratings yet
- Ozdamire 2017organismesDocument7 pagesOzdamire 2017organismesFatma ElzaytNo ratings yet
- Paper Biosis 3Document18 pagesPaper Biosis 3Silvia Estu ZainiNo ratings yet
- Insects 12 00230Document16 pagesInsects 12 0023067kmxfq76rNo ratings yet
- Beetles As Model Organisms in PhysiologiDocument22 pagesBeetles As Model Organisms in Physiologidipeshsingh563No ratings yet
- Research On Orchid Biology and Biotechnology: Plant and Cell Physiology July 2011Document21 pagesResearch On Orchid Biology and Biotechnology: Plant and Cell Physiology July 2011Tyler Jordan DavisonNo ratings yet
- JER 39 (4) Published - Beetles and Forensic-Madhu Bala and Neha Singh - 1-1-2016 PDFDocument11 pagesJER 39 (4) Published - Beetles and Forensic-Madhu Bala and Neha Singh - 1-1-2016 PDFYanna RizkiaNo ratings yet
- Fungal Functional Ecology MSPreprintDocument88 pagesFungal Functional Ecology MSPreprintCamila GuzmanNo ratings yet
- 2012sydowia 64 1 - KlomklungDocument19 pages2012sydowia 64 1 - Klomklung伯尼Bulti Bonsa WondimuNo ratings yet
- Bunga Aulia - A22120062 - Analisis JurnalDocument4 pagesBunga Aulia - A22120062 - Analisis JurnalBunga AuliaNo ratings yet
- tmpF1FC TMPDocument11 pagestmpF1FC TMPFrontiersNo ratings yet
- Composition and Genetic Diversity of Mosquitoes DiDocument17 pagesComposition and Genetic Diversity of Mosquitoes DiNhe FirmansyahNo ratings yet
- CHROMOSOMES STAINING WITH GIEMSA METHODSDocument24 pagesCHROMOSOMES STAINING WITH GIEMSA METHODSNurul HidayahNo ratings yet
- Morphological Characteristics and Phylogenetic AnaDocument15 pagesMorphological Characteristics and Phylogenetic Anamanojtbgri5793No ratings yet
- Progress and Prospects in Rat Genetics: A Community ViewDocument8 pagesProgress and Prospects in Rat Genetics: A Community View454e75376rfufjcbNo ratings yet
- 18066-Article Text-33641-1-10-20181206 PDFDocument10 pages18066-Article Text-33641-1-10-20181206 PDFYasendi AnakriNo ratings yet
- Research in ProtozoologyFrom EverandResearch in ProtozoologyTze-Tuan ChenNo ratings yet
- Effect of PH and Temperature On The Activities of Alphaamylase in Cassava Starch LiquefactionDocument7 pagesEffect of PH and Temperature On The Activities of Alphaamylase in Cassava Starch LiquefactionWelinda MemeNo ratings yet
- Study of Single and Double Sampling PlansDocument14 pagesStudy of Single and Double Sampling PlansTravis WoodNo ratings yet
- ISO 7973 Viscosity BrabenderDocument6 pagesISO 7973 Viscosity BrabenderWelinda MemeNo ratings yet
- Christmas SolicitationDocument2 pagesChristmas SolicitationWelinda Meme100% (1)
- General Principles of Food Hygiene CXC 1-1969Document35 pagesGeneral Principles of Food Hygiene CXC 1-1969AnnaNo ratings yet
- Eurofins Allergen Webinar - 6.2.2021 For Catalyst Consulting IndonesiaDocument27 pagesEurofins Allergen Webinar - 6.2.2021 For Catalyst Consulting IndonesiaWelinda MemeNo ratings yet
- Thermogravimetric and Kinetic Analysis of Cassava Starch Based BioplasticDocument9 pagesThermogravimetric and Kinetic Analysis of Cassava Starch Based BioplasticJoriz FranciscoNo ratings yet
- (Doi 10.1533 - 9781845695750.2.157) Park, Kwan-Hwa - Carbohydrate-Active Enzymes - Enzymatic Modification of Starch For Food IndustryDocument27 pages(Doi 10.1533 - 9781845695750.2.157) Park, Kwan-Hwa - Carbohydrate-Active Enzymes - Enzymatic Modification of Starch For Food IndustryWelinda MemeNo ratings yet
- Dokumen - Tips - Iso Ts 22002 1 PRP Food Safety Food ManufacturingpdfDocument26 pagesDokumen - Tips - Iso Ts 22002 1 PRP Food Safety Food ManufacturingpdfWelinda MemeNo ratings yet
- Water University:: Addressing The World'S Water Challenges Through Science, Technology, Training & DevelopmentDocument4 pagesWater University:: Addressing The World'S Water Challenges Through Science, Technology, Training & DevelopmentWelinda MemeNo ratings yet
- Foods: Relationship Between Water Activity and Moisture Content in Floral HoneyDocument18 pagesFoods: Relationship Between Water Activity and Moisture Content in Floral HoneyWelinda MemeNo ratings yet
- 8476 29366 2 PBDocument13 pages8476 29366 2 PBWelinda MemeNo ratings yet
- The Measurement of Equilibrium Relative Humidity: (Wo WeDocument16 pagesThe Measurement of Equilibrium Relative Humidity: (Wo WeWelinda MemeNo ratings yet
- Catalytic Fast Pyrolysis of Lignocellulosic Biomass: Chemical Society Reviews May 2014Document31 pagesCatalytic Fast Pyrolysis of Lignocellulosic Biomass: Chemical Society Reviews May 2014Welinda MemeNo ratings yet
- Effects of Moisture Content and Storage Period On Proximate Composition, Microbial Counts and Total Carotenoids of Cassava FlourDocument12 pagesEffects of Moisture Content and Storage Period On Proximate Composition, Microbial Counts and Total Carotenoids of Cassava FlourWelinda MemeNo ratings yet
- Guidance Note 18 Determination of Product Shelf Life PDFDocument56 pagesGuidance Note 18 Determination of Product Shelf Life PDFHiếu LêNo ratings yet
- Water University:: Addressing The World'S Water Challenges Through Science, Technology, Training & DevelopmentDocument4 pagesWater University:: Addressing The World'S Water Challenges Through Science, Technology, Training & DevelopmentWelinda MemeNo ratings yet
- Bhakti Trader Ram Pal JiDocument232 pagesBhakti Trader Ram Pal JiplancosterNo ratings yet
- Oracle Apps Quality ModuleDocument17 pagesOracle Apps Quality ModuleSantOsh100% (2)
- District Wise List of Colleges Under The Juridiction of MRSPTU BathindaDocument13 pagesDistrict Wise List of Colleges Under The Juridiction of MRSPTU BathindaGurpreet SandhuNo ratings yet
- Airs-Lms - Math-10 - q3 - Week 3-4 Module 3 Rhonavi MasangkayDocument19 pagesAirs-Lms - Math-10 - q3 - Week 3-4 Module 3 Rhonavi MasangkayRamil J. Merculio100% (1)
- Lesson Plan 2 Revised - Morgan LegrandDocument19 pagesLesson Plan 2 Revised - Morgan Legrandapi-540805523No ratings yet
- Hanwha Engineering & Construction - Brochure - enDocument48 pagesHanwha Engineering & Construction - Brochure - enAnthony GeorgeNo ratings yet
- Presentation 1Document20 pagesPresentation 1nikitakhanduja1304No ratings yet
- Banu Maaruf of The LevantDocument6 pagesBanu Maaruf of The LevantMotiwala AbbasNo ratings yet
- Apps Tables Excel FRMTDocument4 pagesApps Tables Excel FRMTSunil ReddyNo ratings yet
- 5.a Personal Diet Consultant For Healthy MealDocument5 pages5.a Personal Diet Consultant For Healthy MealKishore SahaNo ratings yet
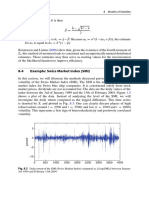
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- Variables in Language Teaching - The Role of The TeacherDocument34 pagesVariables in Language Teaching - The Role of The TeacherFatin AqilahNo ratings yet
- Scientology Abridged Dictionary 1973Document21 pagesScientology Abridged Dictionary 1973Cristiano Manzzini100% (2)
- C11 Strategy DevelopmentDocument30 pagesC11 Strategy DevelopmentPARTI KEADILAN RAKYAT NIBONG TEBALNo ratings yet
- Manual Handling Risk Assessment ProcedureDocument6 pagesManual Handling Risk Assessment ProcedureSarfraz RandhawaNo ratings yet
- Mechanical Engineer with Experience in Heavy Mining Machinery MaintenanceDocument1 pageMechanical Engineer with Experience in Heavy Mining Machinery MaintenanceCertified Rabbits LoverNo ratings yet
- Rotational Equilibrium SimulationDocument3 pagesRotational Equilibrium SimulationCamille ManlongatNo ratings yet
- OMR Sheet Has Been Attached at The End: EST Erial ODocument8 pagesOMR Sheet Has Been Attached at The End: EST Erial OSangeeta BansalNo ratings yet
- List of household items for relocationDocument4 pagesList of household items for relocationMADDYNo ratings yet
- British Isles Composition GuideDocument4 pagesBritish Isles Composition GuidesonmatanalizNo ratings yet
- ProjectDocument86 pagesProjectrajuNo ratings yet
- 19 Unpriced BOM, Project & Manpower Plan, CompliancesDocument324 pages19 Unpriced BOM, Project & Manpower Plan, CompliancesAbhay MishraNo ratings yet
- AnhvancDocument108 pagesAnhvancvanchienha7766No ratings yet
- Olympic Ayres Magic LyricsDocument1 pageOlympic Ayres Magic Lyricsjackliddy96No ratings yet
- College of Physical Therapy Produces Skilled ProfessionalsDocument6 pagesCollege of Physical Therapy Produces Skilled ProfessionalsRia Mae Abellar SalvadorNo ratings yet
- Year 10 Maths PlaneDocument62 pagesYear 10 Maths Planehal wangNo ratings yet
- Solutions: Spheres, Cones and CylindersDocument13 pagesSolutions: Spheres, Cones and CylindersKeri-ann MillarNo ratings yet