You might also like
- 17-7 PHDocument48 pages17-7 PHsunsirNo ratings yet
- CP Write-Up PDFDocument36 pagesCP Write-Up PDFManish KaushikNo ratings yet
- Dietary Apricot Kernel Oil Effect on Hypercholesteremic Rat LiverDocument5 pagesDietary Apricot Kernel Oil Effect on Hypercholesteremic Rat LiverGÖKHAN DURMAZNo ratings yet
- Yamabushitake Mushroom (Hericium Erinaceus) Improved Lipid Metabolism in Mice Fed A High-Fat DietDocument8 pagesYamabushitake Mushroom (Hericium Erinaceus) Improved Lipid Metabolism in Mice Fed A High-Fat DietTute SilvaNo ratings yet
- Seminario 4A Regulacion de La GlicemiaDocument6 pagesSeminario 4A Regulacion de La GlicemiaEdol LopezNo ratings yet
- Alleviation Hysteria Laying Dietary Tryptophan: of in Hens WithDocument5 pagesAlleviation Hysteria Laying Dietary Tryptophan: of in Hens WithImelda MertasariNo ratings yet
- Misslbeck1986 PDFDocument6 pagesMisslbeck1986 PDFRoberto GarceteNo ratings yet
- Xu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsDocument6 pagesXu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsPaul SimononNo ratings yet
- The Mechanism of Dietary Cholesterol Effects On Lipids Metabolism in RatsDocument6 pagesThe Mechanism of Dietary Cholesterol Effects On Lipids Metabolism in RatsNurmalita Sari DewiNo ratings yet
- Metabolic Analysis of BALB/c Mice (Mus Musculus) Exposed in High Fat and Protein Diet Using Human Diagnostics Test KitsDocument16 pagesMetabolic Analysis of BALB/c Mice (Mus Musculus) Exposed in High Fat and Protein Diet Using Human Diagnostics Test KitsEmotholy LogicalNo ratings yet
- Nutrient Physiology, Metabolism, and Nutrient-Nutrient InteractionsDocument5 pagesNutrient Physiology, Metabolism, and Nutrient-Nutrient InteractionsPaul SimononNo ratings yet
- Sterol Carrier Protein and Fatty Acid-Binding ProteinDocument7 pagesSterol Carrier Protein and Fatty Acid-Binding ProteinMartin OreNo ratings yet
- Effects of Spirulina Platensis Microalgae On Antioxidant and Anti-Inflammatory Factors in Diabetic RatsDocument15 pagesEffects of Spirulina Platensis Microalgae On Antioxidant and Anti-Inflammatory Factors in Diabetic Ratsapi-702059884No ratings yet
- Biochemj00227 0033Document7 pagesBiochemj00227 0033handoko pocaNo ratings yet
- Jvms 63 337Document3 pagesJvms 63 337Martin WiniarskiNo ratings yet
- Alterations in Myocardial Lipid Metabolism During Lactation in The RatDocument7 pagesAlterations in Myocardial Lipid Metabolism During Lactation in The RatSharkyNo ratings yet
- Molecular Characterisation of Carbohydrate Digestion and Absorption in Equine Small IntestineDocument10 pagesMolecular Characterisation of Carbohydrate Digestion and Absorption in Equine Small IntestineArthur HeviNo ratings yet
- Research Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsDocument9 pagesResearch Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsJoshNo ratings yet
- 1 s2.0 S002203021100213X MainDocument9 pages1 s2.0 S002203021100213X MainJuniClaudia13No ratings yet
- White 1976Document10 pagesWhite 1976poojajain86No ratings yet
- Quercetin KhasiatDocument6 pagesQuercetin KhasiatTantri AlfionitaNo ratings yet
- Ntiana Manshurica PDFDocument7 pagesNtiana Manshurica PDFcoffey-geoNo ratings yet
- Conjugated Linoleic Acid and Atherosclerosis in RabbitsDocument7 pagesConjugated Linoleic Acid and Atherosclerosis in RabbitstukaeNo ratings yet
- Metformina 1992 Nao HiperglicemicoDocument5 pagesMetformina 1992 Nao HiperglicemicoCássio SilvaNo ratings yet
- Nutrient Metabolism-Research CommunicationDocument4 pagesNutrient Metabolism-Research CommunicationDaffa RamadhaniNo ratings yet
- Enzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesDocument5 pagesEnzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesPadmini RajuNo ratings yet
- Effect of Papaya Leaves & Extract on Growth, Immunity & Efficiency of RabbitsDocument28 pagesEffect of Papaya Leaves & Extract on Growth, Immunity & Efficiency of RabbitsLeonila D. NovioNo ratings yet
- Brown Adipose Tissue Triacylglycerol Synthesis in Rats Adapted To A High-Protein, Carbohydrate-Free DietDocument7 pagesBrown Adipose Tissue Triacylglycerol Synthesis in Rats Adapted To A High-Protein, Carbohydrate-Free DietPaul SimononNo ratings yet
- Dr. Blanco ArticleDocument11 pagesDr. Blanco ArticleCarlosNo ratings yet
- CapsaicinDocument12 pagesCapsaicinExlonk Gil PeláezNo ratings yet
- The Effect of Chitosan on Lipid Profiles in Rat Blood SerumDocument8 pagesThe Effect of Chitosan on Lipid Profiles in Rat Blood SerumWulandariNo ratings yet
- Chlorogenic Acid Improves Obesity and Lipid MetabolismDocument7 pagesChlorogenic Acid Improves Obesity and Lipid MetabolismproluvieslacusNo ratings yet
- Role of Sortilin 1 (SORT1) On Lipid Metabolism in Bovine LiverDocument15 pagesRole of Sortilin 1 (SORT1) On Lipid Metabolism in Bovine Liversanti nurdinNo ratings yet
- Anti Diabetic Effects of Pumpkin and Its Components Trigonelline and Nicotinic Acid On Goto Kakizaki RatsDocument10 pagesAnti Diabetic Effects of Pumpkin and Its Components Trigonelline and Nicotinic Acid On Goto Kakizaki RatsnurrahmahidayatiNo ratings yet
- Rice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityDocument10 pagesRice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityXavier Alvaro Quille MamaniNo ratings yet
- Protective Effect of Tryptophan and Cysteine Against Carbon Tetrachloride-Induced Liver Injury'Document11 pagesProtective Effect of Tryptophan and Cysteine Against Carbon Tetrachloride-Induced Liver Injury'DoomNo ratings yet
- Protein Kinase Cis Activated in From Rats: Glomeruli DiabeticDocument9 pagesProtein Kinase Cis Activated in From Rats: Glomeruli Diabeticapi-19757116No ratings yet
- Sun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDocument7 pagesSun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDaniela Patricia Alvarez AravenaNo ratings yet
- Artigo AntônioDocument17 pagesArtigo AntônioHenry Daniel Ruiz AlbaNo ratings yet
- International Journal of Pharma and Bio Sciences: Research ArticleDocument7 pagesInternational Journal of Pharma and Bio Sciences: Research ArticleShivani ReddyNo ratings yet
- Journal of Global Pharma TechnologyDocument6 pagesJournal of Global Pharma TechnologyBassam Yahay IbraheemNo ratings yet
- Whey Transaminaze PDFDocument12 pagesWhey Transaminaze PDFwwwglroNo ratings yet
- Huang 2013Document10 pagesHuang 2013Stefania DavidNo ratings yet
- FoodChem 2018 JadanDocument28 pagesFoodChem 2018 JadanGaby Garcia GuerreroNo ratings yet
- Collagen DipeptidesDocument9 pagesCollagen DipeptidesMaung MaungNo ratings yet
- Effect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsDocument7 pagesEffect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsvinayNo ratings yet
- Berberine Moderates Glucose and Lipid Metabolism TDocument10 pagesBerberine Moderates Glucose and Lipid Metabolism THitesh VermaNo ratings yet
- The Effect of Gambir's Catechin Isolate on AST and ALT Levels in Male Rats Fed a High Fat DietDocument7 pagesThe Effect of Gambir's Catechin Isolate on AST and ALT Levels in Male Rats Fed a High Fat DietSyahira AlmunNo ratings yet
- Reliability of Molecular Weight Determinations Using SDS-PAGEDocument7 pagesReliability of Molecular Weight Determinations Using SDS-PAGEEsaú Bojorquez VelazquezNo ratings yet
- Mitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingDocument7 pagesMitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingRoy WilsonNo ratings yet
- Pengzhan2003 Article PolysaccharidesFromUlvaPertusaDocument8 pagesPengzhan2003 Article PolysaccharidesFromUlvaPertusaNorberto OcañaNo ratings yet
- How Noscapine Metabolise Heme?Document15 pagesHow Noscapine Metabolise Heme?Chief editorNo ratings yet
- Autolytic Degradation of Chicken Intestinal Proteins: S.N. Jamdar, P. HarikumarDocument9 pagesAutolytic Degradation of Chicken Intestinal Proteins: S.N. Jamdar, P. HarikumarAmalia AnggreiniNo ratings yet
- Changes of Lipid Metabolism in Plasma, Liver and Bile During Cholesterol Gallstone Formation in Rabbit ModelDocument3 pagesChanges of Lipid Metabolism in Plasma, Liver and Bile During Cholesterol Gallstone Formation in Rabbit ModelSaylor Huancollo VilcaNo ratings yet
- Glycosylation of CHO-Derived Recombinant tPA Produced Under Elevated pCODocument7 pagesGlycosylation of CHO-Derived Recombinant tPA Produced Under Elevated pCOСтепан РемыгаNo ratings yet
- Re-Evaluation of Total Sulphur Amino Acid Requirement and Determination of Replacement Value of Cystine For Methionine in Semi-Purified Diets of Juvenile Nile Tilapia, Oreochromis NiloticusDocument7 pagesRe-Evaluation of Total Sulphur Amino Acid Requirement and Determination of Replacement Value of Cystine For Methionine in Semi-Purified Diets of Juvenile Nile Tilapia, Oreochromis Niloticus陈浩No ratings yet
- (+) - Catechin & Proanthocyanidin Fraction of Uncaria Gambir Roxb FiaDocument8 pages(+) - Catechin & Proanthocyanidin Fraction of Uncaria Gambir Roxb FiaiforaNo ratings yet
- 111-118 (15) PJZ-1878-14 13-12-14 Effects of Phytosterols On Growth Performance and Fa - PDFDocument9 pages111-118 (15) PJZ-1878-14 13-12-14 Effects of Phytosterols On Growth Performance and Fa - PDFHussain Ahmad QadriNo ratings yet
- colarma,+WRS 28-4 12161Document11 pagescolarma,+WRS 28-4 12161Luidi MarquesNo ratings yet
- Fast Facts: Long-Chain Fatty Acid Oxidation Disorders: Understand, identify and supportFrom EverandFast Facts: Long-Chain Fatty Acid Oxidation Disorders: Understand, identify and supportNo ratings yet
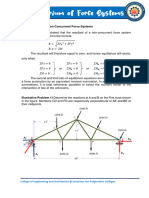
- Chapter 4. Equilibrium of Non Concurrent Force SystemsDocument7 pagesChapter 4. Equilibrium of Non Concurrent Force SystemscarminavegamariaNo ratings yet
- Raman Id To IgDocument8 pagesRaman Id To Igwora123potNo ratings yet
- Bioorganic & Medicinal Chemistry Letters: Janggyoo Choi, Kee Dong Yoon, Jinwoong KimDocument6 pagesBioorganic & Medicinal Chemistry Letters: Janggyoo Choi, Kee Dong Yoon, Jinwoong KimQuesito WistarNo ratings yet
- Corona & Plasma: For ExtrusionDocument8 pagesCorona & Plasma: For ExtrusionHuy Tuan QuachNo ratings yet
- Adminbiotek,+44 File+Utama+Naskah 73 1 11 20210409+Document7 pagesAdminbiotek,+44 File+Utama+Naskah 73 1 11 20210409+Vanesse BarrientosNo ratings yet
- Biomolecules Question Bank (Subjective)Document10 pagesBiomolecules Question Bank (Subjective)Rajendra SahaNo ratings yet
- Monografia MainDocument20 pagesMonografia MainWILIAM SALVADORNo ratings yet
- ChromiumDocument4 pagesChromiumTom SinghNo ratings yet
- Biology Experiments by Vasumitra GajbhiyeDocument18 pagesBiology Experiments by Vasumitra Gajbhiyeinspection100% (1)
- Loctite 380 Black - Ficha TecnicaDocument3 pagesLoctite 380 Black - Ficha TecnicamonsepackNo ratings yet
- Msds Chemguard 3% Afff C-301msDocument4 pagesMsds Chemguard 3% Afff C-301msMatías YaegerNo ratings yet
- Encyclopedia of Electrochemistry, Volume 7b: Inorganic Electrochemistry (Encyclopedia of Electrochemistry) (PDFDrive)Document523 pagesEncyclopedia of Electrochemistry, Volume 7b: Inorganic Electrochemistry (Encyclopedia of Electrochemistry) (PDFDrive)abdulqudus abdulakeemNo ratings yet
- Chem 3Document4 pagesChem 3Cristyl Vismanos GastaNo ratings yet
- (283.15 To 353.15) K at Pressures Up To 25 MP A: PGT Measurements and Eos Predictions of Glycol Ethers FromDocument6 pages(283.15 To 353.15) K at Pressures Up To 25 MP A: PGT Measurements and Eos Predictions of Glycol Ethers FromCarlos FerreraNo ratings yet
- Physical Science SHS 4.3 Worksheet 2Document3 pagesPhysical Science SHS 4.3 Worksheet 2Maricris Jane PeranteNo ratings yet
- Enthalpy change of alcoholsDocument4 pagesEnthalpy change of alcoholsJulia QistinaNo ratings yet
- ProjectDocument12 pagesProjectD E V I LNo ratings yet
- CHEM 43.1 Exercise 5Document5 pagesCHEM 43.1 Exercise 5paradoxcomplexNo ratings yet
- Angus - Case - Ex20.003-E4 Dmmopa TDS 092519Document2 pagesAngus - Case - Ex20.003-E4 Dmmopa TDS 092519Mauro Di FraiaNo ratings yet
- Group 6 - Lab Experiment PhypharmDocument7 pagesGroup 6 - Lab Experiment PhypharmNichole Win MallariNo ratings yet
- Animal Nutrition: Dervan D.S.L. Bryan, Dawn A. Abbott, Henry L. ClassenDocument9 pagesAnimal Nutrition: Dervan D.S.L. Bryan, Dawn A. Abbott, Henry L. ClassenmiminNo ratings yet
- Some Basic Concept of Chemistry QuesDocument40 pagesSome Basic Concept of Chemistry QuesSaadiya HussainNo ratings yet
- Bmac4E02: Modern Analytical InstrumentationDocument22 pagesBmac4E02: Modern Analytical InstrumentationSachin ashokNo ratings yet
- Talanta: Rosy, Himanshu Chasta, Rajendra N. GoyalDocument7 pagesTalanta: Rosy, Himanshu Chasta, Rajendra N. GoyalLeau Sorina-AlexandraNo ratings yet
- Pharmaceutics 13 01313Document23 pagesPharmaceutics 13 01313maizhafiraNo ratings yet
- Kinetics of The Synthesis of Bisphenol A: Applied Catalysis, 37Document10 pagesKinetics of The Synthesis of Bisphenol A: Applied Catalysis, 37khalid -No ratings yet
- Book of PaceDocument207 pagesBook of PaceklikzbypeakyNo ratings yet
- Individual Turnkey Steam Solutions: Pressure Reducing StationsDocument6 pagesIndividual Turnkey Steam Solutions: Pressure Reducing Stationscamilo diazNo ratings yet