You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Raz Lj50 Covidbirthday CLR DsDocument10 pagesRaz Lj50 Covidbirthday CLR DsRocío Bustos SaguaNo ratings yet
- Cognitive Behavioral Treatment For Depressed AdolescentsDocument17 pagesCognitive Behavioral Treatment For Depressed Adolescentscharoline gracetiani nataliaNo ratings yet
- Neonatal BCG Vaccination and Child Survival in TB Exposed and TB Unexposed Children A Prospective Cohort StudyDocument10 pagesNeonatal BCG Vaccination and Child Survival in TB Exposed and TB Unexposed Children A Prospective Cohort Studycharoline gracetiani nataliaNo ratings yet
- Febrile SeizureDocument7 pagesFebrile Seizurecharoline gracetiani nataliaNo ratings yet
- Small Vessel Vasculitides: Key PointsDocument9 pagesSmall Vessel Vasculitides: Key Pointscharoline gracetiani nataliaNo ratings yet
- Relapse in Anorexia NervosaDocument12 pagesRelapse in Anorexia Nervosacharoline gracetiani nataliaNo ratings yet
- Probiotic Allergic RhinitsDocument7 pagesProbiotic Allergic Rhinitscharoline gracetiani nataliaNo ratings yet
- Comparison of Medical Versus Surgical Management of Peritonsillar Abscess: A Retrospective Observational StudyDocument6 pagesComparison of Medical Versus Surgical Management of Peritonsillar Abscess: A Retrospective Observational Studycharoline gracetiani nataliaNo ratings yet
- Chen 2017Document4 pagesChen 2017charoline gracetiani nataliaNo ratings yet
- Academic Anxiety SourcesDocument5 pagesAcademic Anxiety Sourcescharoline gracetiani nataliaNo ratings yet
- Ashoka's Skin DiseaseDocument4 pagesAshoka's Skin DiseaseLalit MishraNo ratings yet
- CNS InfectionDocument36 pagesCNS InfectionIkaDian PuspitanzaNo ratings yet
- Group PolicyDocument14 pagesGroup PolicyMahesh DivakarNo ratings yet
- Case Presentation-Neonatal SepsisDocument36 pagesCase Presentation-Neonatal SepsisDianne DawnNo ratings yet
- Florence College of Nursing Medical Surgical Nursing: Health Talk ONDocument13 pagesFlorence College of Nursing Medical Surgical Nursing: Health Talk ONSunil Kumar100% (1)
- Providing Safe Food and The Phillipine Food Safety FrameworkDocument38 pagesProviding Safe Food and The Phillipine Food Safety Frameworkjanelleallab15No ratings yet
- TB HX Consultation Form A4Document2 pagesTB HX Consultation Form A4Gian Rei MangcucangNo ratings yet
- Cogan Syndrome Surger 4, AnkitDocument18 pagesCogan Syndrome Surger 4, AnkitAnkit Tonger AnkyNo ratings yet
- Study of Cancer: By: Parth Mishra XII-ScienceDocument30 pagesStudy of Cancer: By: Parth Mishra XII-ScienceParth Mishra100% (1)
- CHN REVIEWER LESSON 1 and 2Document9 pagesCHN REVIEWER LESSON 1 and 2Imogen MasumiNo ratings yet
- Questionnaire Pesticide ToxidromeDocument6 pagesQuestionnaire Pesticide ToxidromeKrista Mae LiboNo ratings yet
- Recognizing, Diagnosing, and Treating Mitochondrial DiseaseDocument57 pagesRecognizing, Diagnosing, and Treating Mitochondrial DiseaseSnezana MihajlovicNo ratings yet
- Septicemia PresentationDocument23 pagesSepticemia Presentationapi-548307464No ratings yet
- Gambaran Klinis Dan Tatalaksana SepsisDocument35 pagesGambaran Klinis Dan Tatalaksana SepsisyunielsyaNo ratings yet
- Rnpedia MCN Pnle NaDocument30 pagesRnpedia MCN Pnle NaFrances Sofia DuranNo ratings yet
- Normocytic AnemiaDocument24 pagesNormocytic Anemiasalmocabdinour121No ratings yet
- Sakit Maret 22Document37 pagesSakit Maret 22PKM sukaraja nubanNo ratings yet
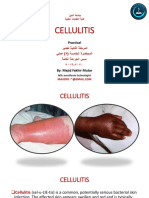
- Cellulitis: PracticalDocument11 pagesCellulitis: PracticalAli dhyaaNo ratings yet
- Theology of DepressionDocument10 pagesTheology of DepressionHubert SelormeyNo ratings yet
- HIV Infection / AIDS (Z21, B20-B24)Document37 pagesHIV Infection / AIDS (Z21, B20-B24)Shreya ThackerNo ratings yet
- Assessment of The Chest and Lungs HandoutDocument2 pagesAssessment of The Chest and Lungs Handoutkaren solibaNo ratings yet
- The Heart 2017Document5 pagesThe Heart 2017Chowdhury Mohammed Tawhid TasneefNo ratings yet
- Case Report: Giant Condyloma Acuminatum of Vulva in An HIV-Infected WomanDocument4 pagesCase Report: Giant Condyloma Acuminatum of Vulva in An HIV-Infected WomanPutri Intan SolehahNo ratings yet
- Increasing Interest of Mass Communication Media and General Public in The Distribution of Tweets About Mental Diseases (Preprint)Document14 pagesIncreasing Interest of Mass Communication Media and General Public in The Distribution of Tweets About Mental Diseases (Preprint)Edwin Rochmad MendiotaNo ratings yet
- Co-Evolution of Parasites and HostsDocument28 pagesCo-Evolution of Parasites and HostsMegbaruNo ratings yet
- Renal Calculi: Dr.K.Sathish Kumar, MD (Hom) .Document13 pagesRenal Calculi: Dr.K.Sathish Kumar, MD (Hom) .mnr hmcNo ratings yet
- Unit 4 ReadingDocument4 pagesUnit 4 ReadingElvantNo ratings yet
- SpeechDocument2 pagesSpeechAryan SinghNo ratings yet
- Pneumococci MIcro LabDocument52 pagesPneumococci MIcro LabSandy AtefNo ratings yet