You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5814)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- L3 Assessment WorkbookDocument55 pagesL3 Assessment WorkbookasadmalikNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
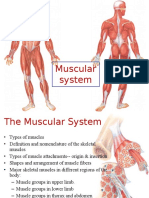
- Muscular SystemDocument26 pagesMuscular Systemlubnab75% (4)
- Fundamentals of Designing Relative Strength ProgramsDocument35 pagesFundamentals of Designing Relative Strength ProgramsFlorian WolfNo ratings yet
- Physiology: by Dr. Mayur SaytaDocument141 pagesPhysiology: by Dr. Mayur Sayta182 ROHIL H100% (1)
- Wearing On Her Nerves Case Study - Werner Williams Mccool Floyd AhmadDocument50 pagesWearing On Her Nerves Case Study - Werner Williams Mccool Floyd Ahmadapi-320365850100% (2)
- JEP Journal of Exercise Physiology: Online OnlineDocument17 pagesJEP Journal of Exercise Physiology: Online OnlineTomas Rivera100% (2)
- Ganong Textbook of Medical PhysiologyDocument98 pagesGanong Textbook of Medical PhysiologyJanie-Vi Gorospe100% (1)
- Resistance Exercise Overtraining and Overreaching: Neuroendocrine ResponsesDocument24 pagesResistance Exercise Overtraining and Overreaching: Neuroendocrine ResponsesAlejandro Albert RequesNo ratings yet
- Mechanisms of Stretch-Induced Muscle Damage in Normal and Dystrophic Muscle: Role of Ionic ChangesDocument13 pagesMechanisms of Stretch-Induced Muscle Damage in Normal and Dystrophic Muscle: Role of Ionic ChangesAlejandro Albert RequesNo ratings yet
- Takarada 2000Document10 pagesTakarada 2000Alejandro Albert RequesNo ratings yet
- Short-Term High-Vs. Low-Velocity Isokinetic Lengthening Training Results in Greater Hypertrophy of The Elbow Exors in Young MenDocument11 pagesShort-Term High-Vs. Low-Velocity Isokinetic Lengthening Training Results in Greater Hypertrophy of The Elbow Exors in Young MenAlejandro Albert RequesNo ratings yet
- Ogborn y Schoenfeld 2017Document25 pagesOgborn y Schoenfeld 2017Alejandro Albert RequesNo ratings yet
- The Underappreciated Role of Muscle in Health and Disease: Review ArticleDocument8 pagesThe Underappreciated Role of Muscle in Health and Disease: Review ArticleAlejandro Albert RequesNo ratings yet
- Emg ProtocolDocument14 pagesEmg ProtocolBrian ThompsonNo ratings yet
- Module 2 - Philippine Meat Industry and Composition of MeatDocument6 pagesModule 2 - Philippine Meat Industry and Composition of MeatVincent Felix A. HernandezNo ratings yet
- DaviesTotal Soccer Fitness PDFDocument167 pagesDaviesTotal Soccer Fitness PDFGaston Saint-Hubert100% (2)
- Biomechanics of MusclesDocument58 pagesBiomechanics of MusclesAsad Chaudhary100% (2)
- Lecture Notes On The Muscular SystemDocument46 pagesLecture Notes On The Muscular SystemDennis Nabor Muñoz, RN,RM100% (1)
- Strength Treatise by C A Sampson PDFDocument250 pagesStrength Treatise by C A Sampson PDFJack100% (1)
- Pex 02 01Document7 pagesPex 02 01QARINA KHAIRUNNISA2010 41No ratings yet
- The Sliding Filament TheoryDocument4 pagesThe Sliding Filament TheoryJinno VeldadNo ratings yet
- Muscle Intelligence 30-Day Primer - Quads & CalvesDocument33 pagesMuscle Intelligence 30-Day Primer - Quads & CalvesPK100% (1)
- Vert Phys Lab Report 1Document3 pagesVert Phys Lab Report 1Erik MingusNo ratings yet
- B.sc. NSG - Sylabus KUHASDocument63 pagesB.sc. NSG - Sylabus KUHASrijumathewmscNo ratings yet
- EasyDocument46 pagesEasyJohnLesterDeLeonNo ratings yet
- Factors That Affect FitnessDocument5 pagesFactors That Affect FitnessStephen San JuanNo ratings yet
- Lucas 1917Document124 pagesLucas 1917abcd_9876No ratings yet
- A&P Ch. 9 NotesDocument5 pagesA&P Ch. 9 Notesslefaveesu6orgNo ratings yet
- CH 10 Lecture PresentationDocument159 pagesCH 10 Lecture PresentationBrian Mathieu Jr.No ratings yet
- Anatomy Ofspinal Cord and Spinal Nerves - ReflexesDocument26 pagesAnatomy Ofspinal Cord and Spinal Nerves - ReflexesDenise MathreNo ratings yet
- 017 Muscle PhysiologyDocument62 pages017 Muscle Physiologyapi-23664998867% (3)
- Physioex 2: Skeletal Muscle PhysiologyDocument3 pagesPhysioex 2: Skeletal Muscle PhysiologyAudreyNo ratings yet
- Muscles and Muscle Tissue: Chapter 9 Part ADocument82 pagesMuscles and Muscle Tissue: Chapter 9 Part ADon KeyNo ratings yet
- Anatomy and Physiology 6th Edition Gunstream Test Bank Full Chapter PDFDocument42 pagesAnatomy and Physiology 6th Edition Gunstream Test Bank Full Chapter PDFserenafinnodx100% (12)
- Skeletal System and Muscular System - REPORTDocument72 pagesSkeletal System and Muscular System - REPORTMhimi ViduyaNo ratings yet
- Utero Toni CosDocument13 pagesUtero Toni CosPedro PachecoNo ratings yet