You might also like
- Clinician S Guide To Neuropsychological AssessmentDocument538 pagesClinician S Guide To Neuropsychological Assessmentmulix8880% (5)
- HHS Public Access: Animal Models of Bone MetastasisDocument38 pagesHHS Public Access: Animal Models of Bone MetastasisDaniela SuarezNo ratings yet
- Vandamme, 2014Document8 pagesVandamme, 2014Khalisha Qintara KhairunnisaNo ratings yet
- Perlman 2016 Mouse Models of Human Disease An EvDocument7 pagesPerlman 2016 Mouse Models of Human Disease An EvChen AiNo ratings yet
- How Evolution Solved Problem of Cancer PreventionDocument5 pagesHow Evolution Solved Problem of Cancer PreventionChristophe MaillardNo ratings yet
- BioEssays - 2022 - Sepp - Wild Animals As An Underused Treasure Trove For Studying The Genetics of CancerDocument5 pagesBioEssays - 2022 - Sepp - Wild Animals As An Underused Treasure Trove For Studying The Genetics of CancerVictor CoutinhoNo ratings yet
- On The Cancer of Wild Animals - Somayeh Zaminpira - Sorush NiknamianDocument6 pagesOn The Cancer of Wild Animals - Somayeh Zaminpira - Sorush NiknamianSomayeh ZaminpiraNo ratings yet
- How Cancer Shapes Evolution, and How Evolution Shapes CancerDocument15 pagesHow Cancer Shapes Evolution, and How Evolution Shapes CancerJoey ZhuNo ratings yet
- Seminars in Immunology: Catharina Hagerling, Zena WerbDocument8 pagesSeminars in Immunology: Catharina Hagerling, Zena WerbdianaNo ratings yet
- Biological Bases and Characterization in OncogenesDocument8 pagesBiological Bases and Characterization in OncogenesCentral Asian StudiesNo ratings yet
- Animal 3Document9 pagesAnimal 3Mehdi MoradiNo ratings yet
- 1081 FullDocument28 pages1081 Fulla khosraviNo ratings yet
- Lowry The Role of Exosomes in Breast CancerDocument9 pagesLowry The Role of Exosomes in Breast CancerCarloz SquallNo ratings yet
- The Cancers That Are Contagious - Discover MagazineDocument7 pagesThe Cancers That Are Contagious - Discover MagazineRoddy PfeifferNo ratings yet
- The Immunobiology of Cancer: An Update ReviewDocument20 pagesThe Immunobiology of Cancer: An Update ReviewicaeeeNo ratings yet
- Esophageal Cancer and Associated GenesDocument16 pagesEsophageal Cancer and Associated GenesEditor IJTSRDNo ratings yet
- Density of Tumor Associated MacrophDocument8 pagesDensity of Tumor Associated MacrophPalloma PortoNo ratings yet
- W de WenisimoDocument18 pagesW de WenisimoBJ CarminatorNo ratings yet
- Cancers Make Their Own Luck - Theories of Cancer OriginsDocument15 pagesCancers Make Their Own Luck - Theories of Cancer Originszhe zhNo ratings yet
- Organo TropismDocument12 pagesOrgano TropismKL TongsonNo ratings yet
- 79 BioTek Ebook FINAL - CompressedDocument63 pages79 BioTek Ebook FINAL - CompressedMichelleNo ratings yet
- Cancer y Celulas Madre 2022Document11 pagesCancer y Celulas Madre 2022Jose EdgarNo ratings yet
- Mariano Marcos State University: College of Health SciencesDocument43 pagesMariano Marcos State University: College of Health ScienceskennethNo ratings yet
- JSCR-18-013Document7 pagesJSCR-18-013Mary ChikamaraNo ratings yet
- Nihms 1768792Document35 pagesNihms 1768792Mohammed KhalidNo ratings yet
- Estrogen and ERα: Potential Culprits in Cervical Cancer DevelopmentDocument17 pagesEstrogen and ERα: Potential Culprits in Cervical Cancer DevelopmentGrace Meigawati IwantoNo ratings yet
- Evolutionary Foundations For Cancer BiologyDocument16 pagesEvolutionary Foundations For Cancer BiologyDenish MikaNo ratings yet
- RussoDocument58 pagesRussoEnvhy WinaNo ratings yet
- Herbs Prevent CancerDocument8 pagesHerbs Prevent CancerErria SuryaNo ratings yet
- p53 My WorkbookDocument12 pagesp53 My WorkbookArunanshu PalNo ratings yet
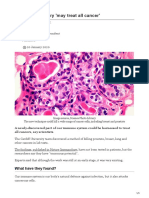
- Immune Discovery May Treat All Cancer - 2020jan20Document5 pagesImmune Discovery May Treat All Cancer - 2020jan20ryan aniceteNo ratings yet
- The Hallmarks of Cancer Review: 2000 by Cell PressDocument14 pagesThe Hallmarks of Cancer Review: 2000 by Cell PressAashiq HussainNo ratings yet
- What is cancerDocument2 pagesWhat is cancerDebapriya HazraNo ratings yet
- Dunn RefDocument47 pagesDunn RefFlorencia Chandrika HalimNo ratings yet
- Hallmark of CancerDocument14 pagesHallmark of CanceriinsabatiniNo ratings yet
- Comparison of Cancer Incidence in Domesticated Versus Wild Animals, As The New Insight Into The Cause and Prevention of Cancer in HumansDocument8 pagesComparison of Cancer Incidence in Domesticated Versus Wild Animals, As The New Insight Into The Cause and Prevention of Cancer in HumansSomayeh ZaminpiraNo ratings yet
- The Cancer HandbookDocument1,529 pagesThe Cancer HandbookAsimaJulianaSiregarNo ratings yet
- PG Art 34580-10Document15 pagesPG Art 34580-10Horia MineaNo ratings yet
- Research Article: Gopinath P.G ., Anitha V.R ., Aruna Mastani SDocument5 pagesResearch Article: Gopinath P.G ., Anitha V.R ., Aruna Mastani SGautami SumanNo ratings yet
- Eva 12018Document9 pagesEva 12018sgutierNo ratings yet
- Piis2405803319302031 PDFDocument4 pagesPiis2405803319302031 PDFRamesh SarmaNo ratings yet
- Zebra FishDocument552 pagesZebra FishmcvalledorNo ratings yet
- Schistosoma ActualidadDocument5 pagesSchistosoma ActualidadWillian EspinozaNo ratings yet
- Immune-Based Cancer Treatment Shows Promise in MiceDocument2 pagesImmune-Based Cancer Treatment Shows Promise in MicemarkzuckerNo ratings yet
- BnhyDocument2 pagesBnhybloozy freezyNo ratings yet
- Journal Pbio 3001471Document20 pagesJournal Pbio 3001471Hisham ShahtoutNo ratings yet
- Paper Cancer MamaDocument10 pagesPaper Cancer MamaMichelle HidroboNo ratings yet
- E Magazine GED106 Group1Document6 pagesE Magazine GED106 Group1Luis SamoyNo ratings yet
- Cell Proliferation and Carcinogenesis in The Hamster Cheek Pouch1Document8 pagesCell Proliferation and Carcinogenesis in The Hamster Cheek Pouch1Om PrakashNo ratings yet
- Liberty University BIOL 101 Individual Assignment 3 Complete Solutions Correct Answers KeyDocument3 pagesLiberty University BIOL 101 Individual Assignment 3 Complete Solutions Correct Answers KeyLiberty0% (1)
- Frontiers in Bioscience E4, 2502-2514, June 1, 2012Document13 pagesFrontiers in Bioscience E4, 2502-2514, June 1, 2012ginocolaciccoNo ratings yet
- Stem Cells and Cancer Stem Cells (2015) PDFDocument159 pagesStem Cells and Cancer Stem Cells (2015) PDFdr.yogaNo ratings yet
- Algae A Robust Living Material Against CancerDocument23 pagesAlgae A Robust Living Material Against CancerJuan David Rodriguez HurtadoNo ratings yet
- Interleukins in Cancer Biology: Their Heterogeneous RoleFrom EverandInterleukins in Cancer Biology: Their Heterogeneous RoleRating: 5 out of 5 stars5/5 (1)
- Cancer de Pancreas en RatasDocument8 pagesCancer de Pancreas en RatasAlethia UkweliNo ratings yet
- Research Paper On Cancer GeneticsDocument7 pagesResearch Paper On Cancer Geneticsrtggklrif100% (1)
- Molecular Biology and Evolution of Cancer: From Discovery To ActionDocument7 pagesMolecular Biology and Evolution of Cancer: From Discovery To ActionPaula Stephany Palacios CastilloNo ratings yet
- Cancer Chemoprevention by Resveratrol: in Vitro and in Vivo Studies and The Underlying Mechanisms (Review)Document12 pagesCancer Chemoprevention by Resveratrol: in Vitro and in Vivo Studies and The Underlying Mechanisms (Review)Daniel Juarez Serrano100% (1)
- Cas 13155Document7 pagesCas 13155Dana LebadaNo ratings yet
- Secondary Metabolites As Lead Molecules For Anticancer Therapies: A ReviewDocument18 pagesSecondary Metabolites As Lead Molecules For Anticancer Therapies: A ReviewVinayNo ratings yet
- Akhtar2019TAF-ChIP An Ultra-Low Input Approach For Genome-Wide Chromatin Immunoprecipitation AssayDocument12 pagesAkhtar2019TAF-ChIP An Ultra-Low Input Approach For Genome-Wide Chromatin Immunoprecipitation AssayKito TongHuiNo ratings yet
- 10 1016@j Jprot 2017 04 002Document25 pages10 1016@j Jprot 2017 04 002Kito TongHuiNo ratings yet
- 10 1016@j Jprot 2017 04 002Document25 pages10 1016@j Jprot 2017 04 002Kito TongHuiNo ratings yet
- Kumar, 2020ATAC-seq Identifies Thousands of Extrachromosomal Circular DNA in Cancer and Cell LinesDocument12 pagesKumar, 2020ATAC-seq Identifies Thousands of Extrachromosomal Circular DNA in Cancer and Cell LinesKito TongHuiNo ratings yet
- Takahashi, 2012 5' End-Centered Expression Profiling Using Cap-Analysis Gene Expression and Next-Generation SequencingDocument43 pagesTakahashi, 2012 5' End-Centered Expression Profiling Using Cap-Analysis Gene Expression and Next-Generation SequencingKito TongHuiNo ratings yet
- Villanacci2011the Spectrum of Drug-Related Colitides Important Entities, Though Frequently OverlookedDocument6 pagesVillanacci2011the Spectrum of Drug-Related Colitides Important Entities, Though Frequently OverlookedKito TongHuiNo ratings yet
- Zhang2018transcriptome Sequencing RNA-SeqDocument13 pagesZhang2018transcriptome Sequencing RNA-SeqKito TongHuiNo ratings yet
- Poulain2017NanoCAGE A Method For The Analysis of Coding and Noncoding 5'-Capped TranscriptomesDocument53 pagesPoulain2017NanoCAGE A Method For The Analysis of Coding and Noncoding 5'-Capped TranscriptomesKito TongHuiNo ratings yet
- Carbon-Based Antiviral Nanomaterials Graphene, C-Dots, and Fullerenes. A PerspectiveDocument32 pagesCarbon-Based Antiviral Nanomaterials Graphene, C-Dots, and Fullerenes. A PerspectiveKito TongHuiNo ratings yet
- Transcriptome Profiling Methods and ApplicationsDocument12 pagesTranscriptome Profiling Methods and ApplicationsKito TongHuiNo ratings yet
- Tonolini2013Acute Nonsteroidal Anti-Inflammatory Drug-Induced ColitisDocument4 pagesTonolini2013Acute Nonsteroidal Anti-Inflammatory Drug-Induced ColitisKito TongHuiNo ratings yet
- Trefoil Factor 3 Isolated From Human Breast Milk Downregulates Cytokines (IL8 and IL6) and Promotes Human Beta Defensin (hBD2 and hBD4) Expression in Intestinal Epithelial Cells HT-29Document10 pagesTrefoil Factor 3 Isolated From Human Breast Milk Downregulates Cytokines (IL8 and IL6) and Promotes Human Beta Defensin (hBD2 and hBD4) Expression in Intestinal Epithelial Cells HT-29Kito TongHuiNo ratings yet
- Nonsteroidal Anti-Inflammatory Drug-Induced Colopathy An Uncommon Cause of Positive Immunochemical Faecal Occult Blood Test in The Program For Colorectal Cancer ScreeningDocument4 pagesNonsteroidal Anti-Inflammatory Drug-Induced Colopathy An Uncommon Cause of Positive Immunochemical Faecal Occult Blood Test in The Program For Colorectal Cancer ScreeningKito TongHuiNo ratings yet
- Transposition of Native Chromatin For Fast and Sensitive Epigenomic Profiling of Open Chromatin, Dna-Binding Proteins and Nucleosome PositionDocument8 pagesTransposition of Native Chromatin For Fast and Sensitive Epigenomic Profiling of Open Chromatin, Dna-Binding Proteins and Nucleosome PositionKito TongHuiNo ratings yet
- darnell2012CLIP (Cross-Linking and Immunoprecipitation) Identification of RNAs Bound by A Specific ProteinDocument16 pagesdarnell2012CLIP (Cross-Linking and Immunoprecipitation) Identification of RNAs Bound by A Specific ProteinKito TongHuiNo ratings yet
- An Injectable Dipeptide-Fullerene Supramolecular Hydrogel For Photodynamic Antibacterial TherapyDocument28 pagesAn Injectable Dipeptide-Fullerene Supramolecular Hydrogel For Photodynamic Antibacterial TherapyKito TongHuiNo ratings yet
- gudkov2019 Unmodified hydrated С60 fullerene molecules exhibit antioxidant properties, prevent damage to DNA and proteins induced by reactive oxygen species and protect mice against injuries caused by radiation-induced oxidative stressDocument32 pagesgudkov2019 Unmodified hydrated С60 fullerene molecules exhibit antioxidant properties, prevent damage to DNA and proteins induced by reactive oxygen species and protect mice against injuries caused by radiation-induced oxidative stressKito TongHuiNo ratings yet
- Anti-Inflammatory Effects of Millet Bran Derived-Bound Polyphenols in LPS-induced HT-29 Cell Signaling PathwayDocument13 pagesAnti-Inflammatory Effects of Millet Bran Derived-Bound Polyphenols in LPS-induced HT-29 Cell Signaling PathwayKito TongHuiNo ratings yet
- Baati2012the Prolongation of The Lifespan of Rats by Repeated Oral Administration of (60) FullereneDocument11 pagesBaati2012the Prolongation of The Lifespan of Rats by Repeated Oral Administration of (60) FullereneKito TongHuiNo ratings yet
- Nonsteroidal Antiinflammatory Drugs and ColitisDocument18 pagesNonsteroidal Antiinflammatory Drugs and ColitisKito TongHuiNo ratings yet
- Mammalian Cell Culture ForDocument22 pagesMammalian Cell Culture ForKito TongHuiNo ratings yet
- Differential Cytokine Expression in Human Intestinal Epithelial Cell LinesDocument9 pagesDifferential Cytokine Expression in Human Intestinal Epithelial Cell LinesKito TongHuiNo ratings yet
- Chassaing2014 Dextran Sulfate Sodium (DSS) - Inducedin MiceDocument14 pagesChassaing2014 Dextran Sulfate Sodium (DSS) - Inducedin MiceKito TongHuiNo ratings yet
- C60 in Olive Oil Causes Light-Dependent Toxicity and Does Not Extend Lifespan in MiceDocument13 pagesC60 in Olive Oil Causes Light-Dependent Toxicity and Does Not Extend Lifespan in MiceKito TongHuiNo ratings yet
- Polymers 10 00701 PDFDocument36 pagesPolymers 10 00701 PDFBertha Catalina RostroNo ratings yet
- Eliminating The Capsule-Like Layer To Promote Glucose Uptake For Hyaluronan Production by Engineered Corynebacterium GlutamicumDocument10 pagesEliminating The Capsule-Like Layer To Promote Glucose Uptake For Hyaluronan Production by Engineered Corynebacterium GlutamicumKito TongHuiNo ratings yet
- Naked Mole-Rat Very-High-Molecular-Mass Hyaluronan Exhibits Superior Cytoprotective PropertiesDocument10 pagesNaked Mole-Rat Very-High-Molecular-Mass Hyaluronan Exhibits Superior Cytoprotective PropertiesKito TongHuiNo ratings yet
- Revisiting The Hallmarks of Cancer The Role of HyaluronanDocument11 pagesRevisiting The Hallmarks of Cancer The Role of HyaluronanKito TongHuiNo ratings yet
- Kulaberoglu2019the Material Properties of Naked Mole-Rat HyaluronanDocument14 pagesKulaberoglu2019the Material Properties of Naked Mole-Rat HyaluronanKito TongHuiNo ratings yet
- GastricDocument207 pagesGastricfavo riteNo ratings yet
- Margi KC Amavata 2Document54 pagesMargi KC Amavata 2hetvi.m.patel.1999No ratings yet
- Mechanical Circulatory Assist Devices A Primer ForDocument20 pagesMechanical Circulatory Assist Devices A Primer ForlakshmiNo ratings yet
- Sarcoma SpecialistDocument5 pagesSarcoma SpecialistaminudinrahmanNo ratings yet
- I. Choose The Meaning of The Underlined Words Using Context CluesDocument4 pagesI. Choose The Meaning of The Underlined Words Using Context CluesMikko GomezNo ratings yet
- U - Book-Studies 53no4-Robarge-Passages-Corrected1Mar PDFDocument104 pagesU - Book-Studies 53no4-Robarge-Passages-Corrected1Mar PDFMehmetZekeriyaÖztürkNo ratings yet
- Naso-Alveolar Moulding For Cleft Lip and PalateDocument41 pagesNaso-Alveolar Moulding For Cleft Lip and PalateCristina M. Laureta100% (1)
- Sop-1-Hand-Hygiene NOSIDocument7 pagesSop-1-Hand-Hygiene NOSINosi Aprilia RinaldiNo ratings yet
- Lesson 2 Bacterial Identification and Processing ModuleDocument20 pagesLesson 2 Bacterial Identification and Processing ModuleTinNo ratings yet
- Laboratory Activity Revil Angelica MDocument16 pagesLaboratory Activity Revil Angelica MAngelica Malacay RevilNo ratings yet
- Biotechnology Applications in Hazardous Waste Treatment - DuphaneDocument13 pagesBiotechnology Applications in Hazardous Waste Treatment - DuphaneAlejandro Castro YaruroNo ratings yet
- Gen Bio PDFDocument6 pagesGen Bio PDFGwyne Shenette Karylle NoscalNo ratings yet
- Research Proposal Power Point PresentationDocument24 pagesResearch Proposal Power Point PresentationSithara JayatungaNo ratings yet
- What Is Bronchitis 2Document3 pagesWhat Is Bronchitis 2lea jumawanNo ratings yet
- DNA Fingerprinting Process & ApplicationsDocument53 pagesDNA Fingerprinting Process & ApplicationsmeenakshiNo ratings yet
- Nursing Care for Clients with InfertilityDocument23 pagesNursing Care for Clients with Infertilityastraeax pandaNo ratings yet
- Novi Lestari 1911304150 B3Document3 pagesNovi Lestari 1911304150 B3FarahNo ratings yet
- Blood Bank PDFDocument178 pagesBlood Bank PDFamit ingaleNo ratings yet
- Chemistry SoftDocument12 pagesChemistry SoftHusayn SaifeeNo ratings yet
- Oral Myofunctional Therapy For The Treatment DTMDocument8 pagesOral Myofunctional Therapy For The Treatment DTMmartaNo ratings yet
- Ritickoval Professional Latin For PharmacistsDocument143 pagesRitickoval Professional Latin For PharmacistsÁnh Ngọc PhanNo ratings yet
- Master's Degree in Periodontology: COURSE 2023-2024Document14 pagesMaster's Degree in Periodontology: COURSE 2023-2024Liseth CarreñoNo ratings yet
- Reading Passages - TEST4Document21 pagesReading Passages - TEST4Bilgin AkbabaNo ratings yet
- Hematopoiesis Reading NotesDocument7 pagesHematopoiesis Reading NotesMemeowwNo ratings yet
- Deadly Pathogens Have A Potent Enemy:: ASP ASPDocument2 pagesDeadly Pathogens Have A Potent Enemy:: ASP ASPHosein HashemianNo ratings yet
- National Mediclaim Policy: BrochureDocument3 pagesNational Mediclaim Policy: Brochureshukla8No ratings yet
- Topics: Nasopharyngeal CarcinomaDocument21 pagesTopics: Nasopharyngeal CarcinomaShruthi Y nursingNo ratings yet
- Leah Alesna Reyes Et Al vs. Sisters of Mercy Hospital Et Al - gr130547Document4 pagesLeah Alesna Reyes Et Al vs. Sisters of Mercy Hospital Et Al - gr130547Dani EsequeNo ratings yet
- Health Statistics and EpidemiologyDocument15 pagesHealth Statistics and EpidemiologyDaniela Nicole Manibog ValentinoNo ratings yet