You might also like
- ZXDC48 FB101 Lithium-Ion Battery User Manual V1.0Document43 pagesZXDC48 FB101 Lithium-Ion Battery User Manual V1.0Luis Cruz96% (24)
- Example of Annual Audit Planning Work ProgramDocument2 pagesExample of Annual Audit Planning Work Programrindwa100% (1)
- All I Need Is Love by Klaus Kinski 0394549163 PDFDocument5 pagesAll I Need Is Love by Klaus Kinski 0394549163 PDFFernanda ArzaguetNo ratings yet
- English Vs Romanian SyntaxDocument7 pagesEnglish Vs Romanian SyntaxAna Maria Chirea-Stoica100% (1)
- tmp9855 TMPDocument9 pagestmp9855 TMPFrontiersNo ratings yet
- Protein Aggregates: An Aging Factor Involved in Cell Death: Etienne Maisonneuve, Benjamin Ezraty, and Sam DukanDocument6 pagesProtein Aggregates: An Aging Factor Involved in Cell Death: Etienne Maisonneuve, Benjamin Ezraty, and Sam DukanUrs GeserNo ratings yet
- Novel Bacteria Degrading N-Acylhomoserine Lactones and Their Use As Quenchers of Quorum-Sensing-Regulated Functions of Plant-Pathogenic BacteriaDocument9 pagesNovel Bacteria Degrading N-Acylhomoserine Lactones and Their Use As Quenchers of Quorum-Sensing-Regulated Functions of Plant-Pathogenic BacteriaAce VincentNo ratings yet
- Perfringens On Chicken Feed Media by Lactobacillus Salivarius and Lactobacillus PlantarumDocument5 pagesPerfringens On Chicken Feed Media by Lactobacillus Salivarius and Lactobacillus PlantarumAdnan AzamNo ratings yet
- Characterization of Bacillus CDocument9 pagesCharacterization of Bacillus CAlfan NasrullahNo ratings yet
- Signals in Spoilage of Bean Sprouts Involvement of Bacterial Quorum-SensingDocument11 pagesSignals in Spoilage of Bean Sprouts Involvement of Bacterial Quorum-SensingKothari VishalNo ratings yet
- 3156-2413-2-PB 2.50.26 PM 6.18.51 PMDocument8 pages3156-2413-2-PB 2.50.26 PM 6.18.51 PMSaiful AbdulNo ratings yet
- Yersinia 1Document10 pagesYersinia 1Veronica EugenioNo ratings yet
- Transmission and Persistence of Methicillin-Resistant Staphylococcus Aureus in Milk, Environment, and Workers in Dairy Cattle FarmsDocument6 pagesTransmission and Persistence of Methicillin-Resistant Staphylococcus Aureus in Milk, Environment, and Workers in Dairy Cattle Farmsadimas89No ratings yet
- In Vitro Chromosomal AberrationsDocument15 pagesIn Vitro Chromosomal AberrationsdrlandgeNo ratings yet
- Protein Extract1Document13 pagesProtein Extract1razor0utNo ratings yet
- J Cell Sci-1993-Perou-99-107Document9 pagesJ Cell Sci-1993-Perou-99-107glodovichiNo ratings yet
- Vieira Etal 2017Document8 pagesVieira Etal 2017Manuel ContrerasNo ratings yet
- tmp42AA TMPDocument8 pagestmp42AA TMPFrontiersNo ratings yet
- Ames TestDocument5 pagesAmes TestDeanNo ratings yet
- Leishmania Amazonensis: Xylitol As Inhibitor of Macrophage Infection and Stimulator of Macrophage Nitric Oxide ProductionDocument6 pagesLeishmania Amazonensis: Xylitol As Inhibitor of Macrophage Infection and Stimulator of Macrophage Nitric Oxide ProductionRogério Ramos MarquesNo ratings yet
- 1 s2.0 S0034528806000397 MainDocument6 pages1 s2.0 S0034528806000397 Mainyujiao MaNo ratings yet
- Photodinamic Inactivation of An RNA EnvelopedDocument7 pagesPhotodinamic Inactivation of An RNA EnvelopedJosé Hiram Sánchez GascaNo ratings yet
- Cloning and Heterologous Expression of Hematin-Dependent Catalase Produced by Lactobacillus Plantarum CNRZ 1228Document4 pagesCloning and Heterologous Expression of Hematin-Dependent Catalase Produced by Lactobacillus Plantarum CNRZ 1228RickyAzrofiSamaraNo ratings yet
- Cholesterol-Lowering Effects of A Putative Probiotic Strain Lactobacillus Plantarum EM Isolated From Kimchi PDFDocument30 pagesCholesterol-Lowering Effects of A Putative Probiotic Strain Lactobacillus Plantarum EM Isolated From Kimchi PDFpniramolNo ratings yet
- ActivinDocument12 pagesActivinHanis Kusumawati RahayuNo ratings yet
- Antibacterial Agents That Inhibit Twocomponent Signal Transduction Systems (In Gram Positive Patogenic Bacteria)Document6 pagesAntibacterial Agents That Inhibit Twocomponent Signal Transduction Systems (In Gram Positive Patogenic Bacteria)Natsume HikariNo ratings yet
- The Lactococcus Lactis PanDocument23 pagesThe Lactococcus Lactis PangabrielNo ratings yet
- Selection of Lactobacilli For Chicken Probiotic Adjuncts: M. Garriga, M. Pascual, J.M. Monfort and M. HugasDocument8 pagesSelection of Lactobacilli For Chicken Probiotic Adjuncts: M. Garriga, M. Pascual, J.M. Monfort and M. Hugasaditi_joshee419No ratings yet
- Pseudomonas Aeruginosa Las Quorum Sensing Autoinducer Suppresses Growth and Biofilm Production in Legionella SpeciesDocument6 pagesPseudomonas Aeruginosa Las Quorum Sensing Autoinducer Suppresses Growth and Biofilm Production in Legionella SpeciesEti ApriyantiNo ratings yet
- Biofilme CampylobacterDocument10 pagesBiofilme CampylobacterJoao Paulo LeôncioNo ratings yet
- Produc KillerDocument12 pagesProduc KillerDenys Rivera GuevaraNo ratings yet
- PQ 0403001990Document6 pagesPQ 0403001990Yuwono WibowoNo ratings yet
- Appl. Environ. Microbiol.-2011-Douglas-7365-71Document8 pagesAppl. Environ. Microbiol.-2011-Douglas-7365-71Nathan McCorkleNo ratings yet
- 1 Zarfel2013Document8 pages1 Zarfel2013luismitlvNo ratings yet
- Aeruginosa Strain PAO1 Is Regulated by Quorum Sensing andDocument15 pagesAeruginosa Strain PAO1 Is Regulated by Quorum Sensing andSarah KKCNo ratings yet
- 1 s2.0 S0168160506000195 Main PDFDocument9 pages1 s2.0 S0168160506000195 Main PDFbernard_china_1No ratings yet
- MarxianusDocument12 pagesMarxianusDarina ilykNo ratings yet
- Curva CrescimentoDocument6 pagesCurva CrescimentoCarina MoitaNo ratings yet
- Cholesterol Assimilation by Lactic Acid Bacteria and Bifidobacteria Isolated From The Human GutDocument5 pagesCholesterol Assimilation by Lactic Acid Bacteria and Bifidobacteria Isolated From The Human GutmicrozasterNo ratings yet
- Armando Et Al 2011Document10 pagesArmando Et Al 2011Adrian Melgratti JobsonNo ratings yet
- Rinaudo 2004Document11 pagesRinaudo 2004Ruyu WangNo ratings yet
- IMVicDocument3 pagesIMVicSundus LiaquatNo ratings yet
- 1005 FullDocument5 pages1005 FullSonia RogersNo ratings yet
- Effects of Recombinant Bovine Interleukin-8 (rbIL-8) Treatment On HealthDocument12 pagesEffects of Recombinant Bovine Interleukin-8 (rbIL-8) Treatment On HealthPablo Cesar Gutierrez Del VillarNo ratings yet
- Al Holy2006 PDFDocument12 pagesAl Holy2006 PDFCarlos LopesNo ratings yet
- Chatterjee 2006Document10 pagesChatterjee 2006Yunita MarwahNo ratings yet
- Pan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.Document5 pagesPan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.AfdhalRuslanNo ratings yet
- Staphylococcus Pettenkoferi Sp. Nov., A Novel Coagulase-Negative Staphylococcal Species Isolated From Human Clinical SpecimensDocument6 pagesStaphylococcus Pettenkoferi Sp. Nov., A Novel Coagulase-Negative Staphylococcal Species Isolated From Human Clinical Specimenski_dvmNo ratings yet
- MelitusDocument6 pagesMelituswulan faddhylanNo ratings yet
- Archivos de Medicina Veterinaria - Comparación de Tres Métodos de Diagnóstico de Paratuberculosis Bovina en Rebaños Lecheros inDocument8 pagesArchivos de Medicina Veterinaria - Comparación de Tres Métodos de Diagnóstico de Paratuberculosis Bovina en Rebaños Lecheros inguadialvarezNo ratings yet
- Carme Plumed-Ferrer 2013Document8 pagesCarme Plumed-Ferrer 2013tenchufo93No ratings yet
- Open Veterinary Journal, (2021), Vol. 11 (1) : 112-120: ISSN: 2226-4485 (Print) ISSN: 2218-6050 (Online) DoiDocument9 pagesOpen Veterinary Journal, (2021), Vol. 11 (1) : 112-120: ISSN: 2226-4485 (Print) ISSN: 2218-6050 (Online) DoiVitoria de Sousa MunizNo ratings yet
- Antimicrobial Activity and Characteristics of Bacteriocins Produced by Vaginal LactobacilliDocument7 pagesAntimicrobial Activity and Characteristics of Bacteriocins Produced by Vaginal LactobacilliManohar PattarNo ratings yet
- Immunological Relationships Bacillus Subsp.: Proteins Making Up The Thuringiensis Israelensis Crystalline ToxinDocument6 pagesImmunological Relationships Bacillus Subsp.: Proteins Making Up The Thuringiensis Israelensis Crystalline ToxinSathish KumarNo ratings yet
- The Multistage Process of Carcinogenesis in Human Esophageal Epithelial Cells Induced by Human PapillomavirusDocument8 pagesThe Multistage Process of Carcinogenesis in Human Esophageal Epithelial Cells Induced by Human PapillomavirusIlma Kurnia SariNo ratings yet
- tác hại của thuốc kháng sinh trong thịt gàDocument2 pagestác hại của thuốc kháng sinh trong thịt gàPhước AnNo ratings yet
- Interaction Between P97/valosin-Containing Protein (VCP) and CSN5 in Early Human PlacentaDocument17 pagesInteraction Between P97/valosin-Containing Protein (VCP) and CSN5 in Early Human PlacentadrmacrohardNo ratings yet
- International Journal of Cancer Volume 85 Issue 3 2000Document8 pagesInternational Journal of Cancer Volume 85 Issue 3 2000BenePicarNo ratings yet
- Antitumor Activity of Cratylia Mollis Lectin Encapsulated Into LiposomesDocument11 pagesAntitumor Activity of Cratylia Mollis Lectin Encapsulated Into LiposomesAlbertoNo ratings yet
- Detection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.Document6 pagesDetection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.International Medical PublisherNo ratings yet
- Staphylococcus Aureus, The Most Virulent Staphylococcus Species, Is Also The Prevalent Pathogen IsolatedDocument6 pagesStaphylococcus Aureus, The Most Virulent Staphylococcus Species, Is Also The Prevalent Pathogen IsolatedVivi RocheNo ratings yet
- 16S Nanopore Sequencing of Lactobacillus - SPP in Apis Mellifera, and Investigation of Their Bacteriocin ActivityDocument21 pages16S Nanopore Sequencing of Lactobacillus - SPP in Apis Mellifera, and Investigation of Their Bacteriocin ActivityLaura PerezNo ratings yet
- Pathological and Microbiological Characterization of Mastitis in Dairy CowsDocument10 pagesPathological and Microbiological Characterization of Mastitis in Dairy CowsLatifa Putri FajrNo ratings yet
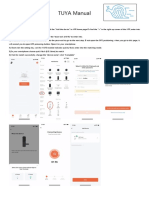
- TUYA ManualDocument2 pagesTUYA ManualMagandang MallNo ratings yet
- Interweave Fiber Catalog Fall 2009Document56 pagesInterweave Fiber Catalog Fall 2009Interweave100% (1)
- Variable Frequency DriveDocument8 pagesVariable Frequency DriveNAYEEM100% (1)
- Universiti Teknologi Mara (Uitm) Shah Alam, MalaysiaDocument4 pagesUniversiti Teknologi Mara (Uitm) Shah Alam, MalaysiaSuraya AdriyanNo ratings yet
- Principles of MicroeconomicsDocument41 pagesPrinciples of MicroeconomicsHoàng LýNo ratings yet
- V Packing SealDocument10 pagesV Packing SealBobby ArbianugrohoNo ratings yet
- Research Title Approval FormDocument2 pagesResearch Title Approval FormJames EdlynNo ratings yet
- Prima Magistra:: Wulan Rahayu Syachtiyani, Novi TrisnawatiDocument12 pagesPrima Magistra:: Wulan Rahayu Syachtiyani, Novi TrisnawatiGita GloriaNo ratings yet
- 3838-Article Text-7786-1-10-20230403Document8 pages3838-Article Text-7786-1-10-20230403Ramona Elena SpiridonNo ratings yet
- Tcexam Result 3174 419 20111208Document8 pagesTcexam Result 3174 419 20111208goldenkumarNo ratings yet
- Tankless Vs TankDocument2 pagesTankless Vs TankClick's PlumbingNo ratings yet
- Electrical Service Minimum RequirmentDocument97 pagesElectrical Service Minimum RequirmentachniajosiNo ratings yet
- Literature Review 1Document7 pagesLiterature Review 1api-609504422No ratings yet
- DSO Digital Storage Oscilloscope: ApplicationDocument2 pagesDSO Digital Storage Oscilloscope: ApplicationmsequipmentsNo ratings yet
- MUMBAI ConsultantsDocument43 pagesMUMBAI ConsultantsER RaviNo ratings yet
- Multidimensional Flow Modeling of The Compression Test of A Gaede Pump Stage in The Viscous RegimeDocument9 pagesMultidimensional Flow Modeling of The Compression Test of A Gaede Pump Stage in The Viscous RegimeAsif HameedNo ratings yet
- Uvm UnderstandingDocument8 pagesUvm UnderstandingLokesh KumarNo ratings yet
- Artikel Jurnal Siti Tsuwaibatul ADocument11 pagesArtikel Jurnal Siti Tsuwaibatul Aaslamiyah1024No ratings yet
- 2020-21 Series Test 1 QPDocument1 page2020-21 Series Test 1 QPred18ggmuNo ratings yet
- Sika Decap PDFDocument2 pagesSika Decap PDFthe pilotNo ratings yet
- Validation of Analytical ProceduresDocument15 pagesValidation of Analytical ProceduresildamonalisaNo ratings yet
- GCE A - AS Level Biology A Topic Test - Biodiversity, Evolution and DiseaseDocument25 pagesGCE A - AS Level Biology A Topic Test - Biodiversity, Evolution and Diseasearfaat shahNo ratings yet
- Proper Storage of Instruments2Document20 pagesProper Storage of Instruments2Pierre Vincent PorrasNo ratings yet
- 12auxiliary ViewsDocument22 pages12auxiliary Viewsmanishjangid9869No ratings yet
- MgF2 SolGelDocument8 pagesMgF2 SolGelumut bayNo ratings yet
- What Is A RubricDocument2 pagesWhat Is A Rubricjasmina2869No ratings yet