You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Mountain Dog Training Program 15 The ExecutionerDocument73 pagesMountain Dog Training Program 15 The ExecutionerMiguel Moreno Ortega100% (5)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Kinematic and Kinetic Differences in Block and Split-Stance Standing Starts During 30 M Sprint-RunningDocument10 pagesKinematic and Kinetic Differences in Block and Split-Stance Standing Starts During 30 M Sprint-Runningok okNo ratings yet
- Selter, P - Assault Bike WOD Bible! - 80 Air Assault Bike Workouts From Hell That'Ll Build Strength, Speed & Endurance (2016)Document71 pagesSelter, P - Assault Bike WOD Bible! - 80 Air Assault Bike Workouts From Hell That'Ll Build Strength, Speed & Endurance (2016)ok okNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Chi Nei Tsangi 2 - Internal Organs Chi Massage PDFDocument66 pagesChi Nei Tsangi 2 - Internal Organs Chi Massage PDFAlexandra Ioana NiculescuNo ratings yet
- Turmiello, Nick - Strength Zone Training The Most Effective Method For Maximizing Muscle Development (2022)Document330 pagesTurmiello, Nick - Strength Zone Training The Most Effective Method For Maximizing Muscle Development (2022)ok ok100% (4)
- Case Study: Mulligan Concept Management of "Tennis Elbow"Document3 pagesCase Study: Mulligan Concept Management of "Tennis Elbow"noonoons100% (1)
- Kinematic and Kinetic Parameters of Sprint Start ADocument11 pagesKinematic and Kinetic Parameters of Sprint Start Aok okNo ratings yet
- 20230331091457-2023-Does Lactate-Guided Threshold Interval Training Within ADocument15 pages20230331091457-2023-Does Lactate-Guided Threshold Interval Training Within ARichardNo ratings yet
- Biomechanical Differences in The Sprint Start Between Faster and Slower High-Level SprintersDocument11 pagesBiomechanical Differences in The Sprint Start Between Faster and Slower High-Level Sprintersok okNo ratings yet
- Davis2021 Article In-SeasonNutritionStrategiesAnDocument23 pagesDavis2021 Article In-SeasonNutritionStrategiesAnok okNo ratings yet
- Saibene Et, Al. 1985 - Energy Sources in Alpine Skiing (Giant Slalom)Document5 pagesSaibene Et, Al. 1985 - Energy Sources in Alpine Skiing (Giant Slalom)ok okNo ratings yet
- By The Final Submission Deadline (ANA Flights:18:00 One Day Prior To Departure)Document2 pagesBy The Final Submission Deadline (ANA Flights:18:00 One Day Prior To Departure)ok okNo ratings yet
- Hopkins WG Et - Al, 2000 - Measures of Reliability in Sports Medicine and ScienceDocument4 pagesHopkins WG Et - Al, 2000 - Measures of Reliability in Sports Medicine and Scienceok okNo ratings yet
- Kitai 1989 Et - Al, - Specificity of Joint Angle in Isometric TrainingDocument5 pagesKitai 1989 Et - Al, - Specificity of Joint Angle in Isometric Trainingok okNo ratings yet
- M S T I S P G: Aximal Trength Raining Mproves Trength Erformance in RapplersDocument7 pagesM S T I S P G: Aximal Trength Raining Mproves Trength Erformance in Rapplersok okNo ratings yet
- Mechanical Output in Jumps of Marmosets (Callithrix Jacchus)Document7 pagesMechanical Output in Jumps of Marmosets (Callithrix Jacchus)ok okNo ratings yet
- J.L Ivy, Et - Al, 1980 - Muscle Respiratory Capacity and Fiber Type As Determinants of The Lactate ThresholdDocument5 pagesJ.L Ivy, Et - Al, 1980 - Muscle Respiratory Capacity and Fiber Type As Determinants of The Lactate Thresholdok okNo ratings yet
- A.Tillin Et - Al, - Short-Term Unilateral Resistance Training Affects The Agonist-Antagonist But Not The Force-Agonist Activation RelationshipDocument10 pagesA.Tillin Et - Al, - Short-Term Unilateral Resistance Training Affects The Agonist-Antagonist But Not The Force-Agonist Activation Relationshipok okNo ratings yet
- The Dose-Response Relationship Between Interval-Training and VO in Well-Trained Endurance Runners: A Systematic ReviewDocument19 pagesThe Dose-Response Relationship Between Interval-Training and VO in Well-Trained Endurance Runners: A Systematic Reviewok okNo ratings yet
- J.Duchateau Et - Al, - Training Adaptations in The Behavior of Human Motor UnitsDocument11 pagesJ.Duchateau Et - Al, - Training Adaptations in The Behavior of Human Motor Unitsok okNo ratings yet
- M.jordan Et Al. 2015 - Lower Lib Asymmetry in Mechanical Muscle Function - A Comparison Between Ski Racers With and Without ACL ReconstructionDocument9 pagesM.jordan Et Al. 2015 - Lower Lib Asymmetry in Mechanical Muscle Function - A Comparison Between Ski Racers With and Without ACL Reconstructionok okNo ratings yet
- Baxter Et - Al, 2020 - Exercise Progression To Incrementally Load The Achilles TendonDocument7 pagesBaxter Et - Al, 2020 - Exercise Progression To Incrementally Load The Achilles Tendonok okNo ratings yet
- Hayes.P Et - Al, 2012 - Foot Strike Patterns and Ground Contact Times During High-Calibre Middle-Distance RacesDocument10 pagesHayes.P Et - Al, 2012 - Foot Strike Patterns and Ground Contact Times During High-Calibre Middle-Distance Racesok okNo ratings yet
- Bertil Sjodin Et - Al, 1982 - Changes in Onset of Blood Lactate Accumulation (OBLA) and Muscle Enzymes After Training at OBLADocument13 pagesBertil Sjodin Et - Al, 1982 - Changes in Onset of Blood Lactate Accumulation (OBLA) and Muscle Enzymes After Training at OBLAok okNo ratings yet
- "Question Your Categories": The Misunderstood Complexity of Middle-Distance Running Profiles With Implications For Research Methods and ApplicationDocument8 pages"Question Your Categories": The Misunderstood Complexity of Middle-Distance Running Profiles With Implications For Research Methods and Applicationok okNo ratings yet
- A.Jamnick.N Et - Al, 2018 - Manipulating Graded Exercise Test Variables Affects The Validity of The Lactate Threshold and V O2peakDocument21 pagesA.Jamnick.N Et - Al, 2018 - Manipulating Graded Exercise Test Variables Affects The Validity of The Lactate Threshold and V O2peakok okNo ratings yet
- Epi Physio Lys IsDocument24 pagesEpi Physio Lys IsVita TantiNo ratings yet
- Elbow Problems: Paralysis: Upper Limb OrthoticsDocument46 pagesElbow Problems: Paralysis: Upper Limb OrthoticsJalil AhmadNo ratings yet
- Saxophone Fingering Chart: All The Fingerings You Need To Get Started Playing The Saxophone in One Handy GuideDocument7 pagesSaxophone Fingering Chart: All The Fingerings You Need To Get Started Playing The Saxophone in One Handy GuideDan AcarNo ratings yet
- Postural DeformitiesDocument37 pagesPostural DeformitiesBalram Jha80% (5)
- Program Design 101 PDFDocument13 pagesProgram Design 101 PDFjole000No ratings yet
- Performance Checklist - ROMDocument3 pagesPerformance Checklist - ROMCrishaGarraNo ratings yet
- Epos Hip UltrasoundDocument20 pagesEpos Hip UltrasoundBogdan Popescu100% (1)
- Posterior Hip Dislocation Case FileDocument2 pagesPosterior Hip Dislocation Case Filehttps://medical-phd.blogspot.comNo ratings yet
- A&P - Muscle WorksheetDocument4 pagesA&P - Muscle WorksheetJerry GNo ratings yet
- Moh Giles 1Document30 pagesMoh Giles 1Divya AggarwalNo ratings yet
- Splinting Pocket Guide PDFDocument16 pagesSplinting Pocket Guide PDFMohd Khairil Anwar Ramli100% (1)
- 2010 A Turner - The Stretch-Shortening Cycle - Proposed Mechanisms and Methods For EnhancementDocument13 pages2010 A Turner - The Stretch-Shortening Cycle - Proposed Mechanisms and Methods For EnhancementLuis VargasNo ratings yet
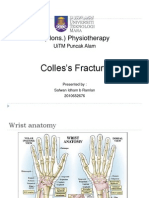
- Colles's Fracture: B (Hons.) PhysiotherapyDocument36 pagesColles's Fracture: B (Hons.) PhysiotherapySafwan Idham RamlanNo ratings yet
- Distal Biceps Tendon InjuriesDocument9 pagesDistal Biceps Tendon InjuriesEllan Giulianno FerreiraNo ratings yet
- Physical Education & Health: Exercies and HomeworkoutDocument21 pagesPhysical Education & Health: Exercies and HomeworkoutMåřïä Ļà ĞŕëàthaNo ratings yet
- Patellofemoral Pain Syndrome (Runner'S Knee)Document2 pagesPatellofemoral Pain Syndrome (Runner'S Knee)Dewi IrfanNo ratings yet
- TMJ Exercises: Specialist SurgeryDocument8 pagesTMJ Exercises: Specialist SurgeryMassimiliano MarchionneNo ratings yet
- Manipulations For TuinaDocument95 pagesManipulations For TuinaFabrizio F. Caragnano100% (2)
- Morning Report: Pembimbing: Dr. Djunifer Hasudungan Sagala, Sp. OT (K) Dr. Andreas Wahyu W, Sp. OTDocument55 pagesMorning Report: Pembimbing: Dr. Djunifer Hasudungan Sagala, Sp. OT (K) Dr. Andreas Wahyu W, Sp. OTSinda AgathaNo ratings yet
- Daftar Pustaka: Ir - Perpustakaan Universitas AirlanggaDocument3 pagesDaftar Pustaka: Ir - Perpustakaan Universitas AirlanggaDeviNo ratings yet
- How To Get The Best Results From Your DumbbellsDocument8 pagesHow To Get The Best Results From Your DumbbellsKenneth LiNo ratings yet
- Physical Therapy Protocols For Ankle and Foot ConditionsDocument116 pagesPhysical Therapy Protocols For Ankle and Foot ConditionstinadasilvaNo ratings yet
- Foam Rolling GuideDocument5 pagesFoam Rolling GuideGeraldyneNo ratings yet
- Kai Triceps Compressed - UpdatedDocument22 pagesKai Triceps Compressed - UpdatedSundararajan JagannathanNo ratings yet
- Fitness Exercise: Movement EnhancementDocument35 pagesFitness Exercise: Movement EnhancementIgnacio Janina Grace100% (6)
- Tabel Rehab Fraktur (Hoppenfeld) PDFDocument2 pagesTabel Rehab Fraktur (Hoppenfeld) PDFJerry Kurnia WahyudiNo ratings yet
- Presentation (1) KinesiologyDocument24 pagesPresentation (1) Kinesiologyمہک جمالیNo ratings yet