You might also like
- Pathogenesis and Classification of Eosinophil Disorders A Review of Recent Developments in The FieldDocument21 pagesPathogenesis and Classification of Eosinophil Disorders A Review of Recent Developments in The FieldpanduNo ratings yet
- Piciorul Diabetic CursDocument59 pagesPiciorul Diabetic Cursmichelle misu100% (1)
- Curs 14 Boala Lyme 2Document63 pagesCurs 14 Boala Lyme 2n1u1s1h1100% (1)
- Integrarea MetabolismuluiDocument29 pagesIntegrarea MetabolismuluiMunteanu DoruNo ratings yet
- Psoriasis Which Therapy For Which PatientDocument11 pagesPsoriasis Which Therapy For Which PatientWenny AgustinNo ratings yet
- Transfer RNA Synthesis RegulationDocument17 pagesTransfer RNA Synthesis RegulationAfif BerlianNo ratings yet
- Colonization, Infection, and The Accessory Genome of KlebsiellaDocument15 pagesColonization, Infection, and The Accessory Genome of KlebsiellaStephanie ZengNo ratings yet
- Infectii CutanateDocument24 pagesInfectii CutanateMaria Minodora PascuNo ratings yet
- Toxicologie - Notiuni IntroductiveDocument15 pagesToxicologie - Notiuni IntroductiveDaniela GreereNo ratings yet
- Acute InflammationDocument10 pagesAcute InflammationLydia Angel HartonoNo ratings yet
- Plasmodium ReportDocument43 pagesPlasmodium ReportGenessa Agustin BuenafeNo ratings yet
- AscarisDocument42 pagesAscarisمصطفي خندقاوي100% (1)
- ToxicologieDocument29 pagesToxicologieRazvan FumeaNo ratings yet
- Immunology Mucosal and Body Surface Andrew e 2165513Document1 pageImmunology Mucosal and Body Surface Andrew e 2165513Ar khaisanNo ratings yet
- Merkel Cell CarcinomaDocument15 pagesMerkel Cell CarcinomaJhonnathan Rodriguez100% (1)
- Psoriasis: Signs, Symptoms, DiagnosisDocument45 pagesPsoriasis: Signs, Symptoms, DiagnosisAnonymous 9Sf9xCMRgfNo ratings yet
- Atopic Dermatitis and Skin Microbiome Ne PDFDocument6 pagesAtopic Dermatitis and Skin Microbiome Ne PDFSmaranda ADNo ratings yet
- LMC CursDocument31 pagesLMC CursEma ȚurcașNo ratings yet
- Inflammation Chronic Diseases and Cancer - Cell and Molecular Biology Immunology and Clinical Bases PDFDocument442 pagesInflammation Chronic Diseases and Cancer - Cell and Molecular Biology Immunology and Clinical Bases PDFsatriomega100% (1)
- 【机制】IL-12 and IL-23 Pathway Inhibition in Inflammatory Bowel DiseaseDocument14 pages【机制】IL-12 and IL-23 Pathway Inhibition in Inflammatory Bowel Diseasejona renNo ratings yet
- Charcot-Marie-Tooth DiseaseDocument9 pagesCharcot-Marie-Tooth Diseaseilyas9558No ratings yet
- Ghid Angelescu 2018 ScanDocument144 pagesGhid Angelescu 2018 ScanAna RotariuNo ratings yet
- HematocritDocument2 pagesHematocritማላያላም ማላያላምNo ratings yet
- EUCAST E Def 7.3.2 Yeast Testing Definitive Revised 2020Document21 pagesEUCAST E Def 7.3.2 Yeast Testing Definitive Revised 2020Guneyden GuneydenNo ratings yet
- Genul StaphylococcusDocument28 pagesGenul StaphylococcusAna Maria RusuNo ratings yet
- The Difference Between Semiotics and SemDocument17 pagesThe Difference Between Semiotics and SemNora Natalia LerenaNo ratings yet
- IHC - InterpretareDocument185 pagesIHC - InterpretareAnca NeaguNo ratings yet
- Anti TGDocument24 pagesAnti TGFlorea RodicaNo ratings yet
- Introduction To Enzyme-Linked Immunosorbent Assay (ELISA)Document34 pagesIntroduction To Enzyme-Linked Immunosorbent Assay (ELISA)Bem LimNo ratings yet
- Diagnosing Paraprotein Diseases with Lab Tests and Bone Marrow ExamsDocument29 pagesDiagnosing Paraprotein Diseases with Lab Tests and Bone Marrow ExamsScarlettMLVNo ratings yet
- Glandele EndocrineDocument5 pagesGlandele EndocrineIrinaNo ratings yet
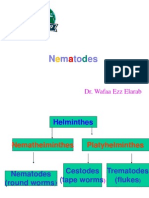
- N M To Es: Dr. Wafaa Ezz ElarabDocument31 pagesN M To Es: Dr. Wafaa Ezz Elarabshoroq zaki100% (1)
- Microbiology: Pathogenic Gram-Positive Cocci (Streptococcus)Document26 pagesMicrobiology: Pathogenic Gram-Positive Cocci (Streptococcus)Shuler0071No ratings yet
- IP PROTOCOL FOR PROTEIN PURIFICATIONDocument6 pagesIP PROTOCOL FOR PROTEIN PURIFICATIONhalfangle100% (1)
- List of Publications (2014-2019Document22 pagesList of Publications (2014-2019Loan LoanNo ratings yet
- Medication Safetyin PolypharmacyDocument64 pagesMedication Safetyin PolypharmacyNestor PerezNo ratings yet
- Probiotics Powerpoint - Lisa G 252c Laura 252c SarahDocument21 pagesProbiotics Powerpoint - Lisa G 252c Laura 252c Sarahapi-322463366No ratings yet
- Culegere de Rezumate - Abstract Book - SRATI 2019Document116 pagesCulegere de Rezumate - Abstract Book - SRATI 2019Mihai PopescuNo ratings yet
- Balasa2020 - Marine Biocompounds For Neuroprotection-A ReviewDocument24 pagesBalasa2020 - Marine Biocompounds For Neuroprotection-A ReviewMark AñazcoNo ratings yet
- Status of Etoricoxib in The Treatment of Rheumatic Diseases. Expert Panel OpinionDocument8 pagesStatus of Etoricoxib in The Treatment of Rheumatic Diseases. Expert Panel OpinionHendri Yanto100% (1)
- ElectrophoresisDocument32 pagesElectrophoresismadhaviNo ratings yet
- Test Questionairre FormatDocument149 pagesTest Questionairre FormatSantosh Kumar TataNo ratings yet
- Prepared By: Murari Pavan M.Pharm (Pharmaceutics) Gautham College of PharmacyDocument22 pagesPrepared By: Murari Pavan M.Pharm (Pharmaceutics) Gautham College of PharmacyAshutosh LohumiNo ratings yet
- Pharmacogenetics and Pharmacogenomics: Kevin Zbuk, MD Medical Oncologist Juravinski Cancer Centre Mcmaster UniversityDocument72 pagesPharmacogenetics and Pharmacogenomics: Kevin Zbuk, MD Medical Oncologist Juravinski Cancer Centre Mcmaster UniversityAnubhuti JhaNo ratings yet
- Dermatology: Dr. N. Shear Blaise Clarkson and Elana Lavine, Chapter Editors Harriette Van Spall, Associate EditorDocument44 pagesDermatology: Dr. N. Shear Blaise Clarkson and Elana Lavine, Chapter Editors Harriette Van Spall, Associate EditorBraga LaviniaNo ratings yet
- Palmoplantar Psoriasis, Palma-Plantar PsoriasisDocument2 pagesPalmoplantar Psoriasis, Palma-Plantar Psoriasisapi-19522927No ratings yet
- Isolation, Identification & Detection of Microbes: Meenakshi Kashyap M.Phil Botany Course 2B: Research Methodology in 2ADocument62 pagesIsolation, Identification & Detection of Microbes: Meenakshi Kashyap M.Phil Botany Course 2B: Research Methodology in 2AmujeebrkmNo ratings yet
- Charcot Foot - An Update PDFDocument6 pagesCharcot Foot - An Update PDFgamalramadiNo ratings yet
- Arsene Lupin, Gentleman Burglar - Maurice LeblancDocument174 pagesArsene Lupin, Gentleman Burglar - Maurice LeblancIan VíctorNo ratings yet
- Left Testicular Vein (SN)Document1 pageLeft Testicular Vein (SN)Joyabrata SarkarNo ratings yet
- The Use of Latin Terminology in Medical Case Reports Quantitative, Structural, and Thematic AnalysisDocument10 pagesThe Use of Latin Terminology in Medical Case Reports Quantitative, Structural, and Thematic Analysisbose_lowe11No ratings yet
- MRSADocument44 pagesMRSAMarion PerniaNo ratings yet
- DR Ara Khan Marwat: Join My Watssap Group For Part-1 Books, Papers and UpdatesDocument25 pagesDR Ara Khan Marwat: Join My Watssap Group For Part-1 Books, Papers and UpdatesmisdduaaNo ratings yet
- Blood Test ResultsDocument1 pageBlood Test ResultsnindyaNo ratings yet
- T.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigDocument18 pagesT.G.J. Allen and G. Burnstock - Intracellular Studies of The Electrophysiological Properties of Cultured Intracardiac Neurones of The Guinea-PigLuammmNo ratings yet
- Histological Basis of The Porcine Femoral Artery For Vascular ResearchDocument7 pagesHistological Basis of The Porcine Femoral Artery For Vascular ResearchaneliatiarasuciNo ratings yet
- 543 FullDocument6 pages543 FullantoniorNo ratings yet
- TMP AE68Document13 pagesTMP AE68FrontiersNo ratings yet
- Temporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary RunningDocument8 pagesTemporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary Runningsonjeonggyu87No ratings yet
- Telemetry: C-Fos and Dfosb Immunoreactivity in Rat Brain JT Cunningham Et AlDocument2 pagesTelemetry: C-Fos and Dfosb Immunoreactivity in Rat Brain JT Cunningham Et AlMicheal ChungNo ratings yet
- Clinical Lab TechnqDocument67 pagesClinical Lab TechnqRatnamala SarmaNo ratings yet
- Science 5 Quiz Bee ReviewerDocument7 pagesScience 5 Quiz Bee ReviewerRommel Urbano Yabis50% (2)
- Applied Mechanics - 341-345 NSQF IrdtDocument4 pagesApplied Mechanics - 341-345 NSQF IrdtTushar KiranNo ratings yet
- Notes in Readings in Philippine History: Act of Proclamation of Independence of The Filipino PeopleDocument4 pagesNotes in Readings in Philippine History: Act of Proclamation of Independence of The Filipino PeopleAiris Ramos AgillonNo ratings yet
- Labour Law Course WorkDocument5 pagesLabour Law Course WorkUncle BrianNo ratings yet
- Learn Basic Music Notes in Grade 3Document3 pagesLearn Basic Music Notes in Grade 3Ned Banania PagunsanNo ratings yet
- Choosing Ayurveda As A Healthcare PractiDocument16 pagesChoosing Ayurveda As A Healthcare PractiAndie PebblesNo ratings yet
- Hybrid - 01Document209 pagesHybrid - 01Santos ArwinNo ratings yet
- Tijing Vs CA DigestDocument1 pageTijing Vs CA DigestLeo GuillermoNo ratings yet
- OMCC Board ResolutionDocument1 pageOMCC Board ResolutionOrient MansionNo ratings yet
- Din Ling, 'Miss Sophia's Diary'Document2 pagesDin Ling, 'Miss Sophia's Diary'StudenteNo ratings yet
- UNIT 4 Microwave TubesDocument39 pagesUNIT 4 Microwave TubesSnigdha SidduNo ratings yet
- Reading Comprehension Worksheets For Grade 5Document15 pagesReading Comprehension Worksheets For Grade 5Lhey Marie Arroz PedrosNo ratings yet
- English Proficiency TrainingDocument21 pagesEnglish Proficiency TrainingDayang GNo ratings yet
- Caste and Class An Interlinked ViewDocument12 pagesCaste and Class An Interlinked ViewghisaramNo ratings yet
- A. Two Subsequences: Codeforces Round #751 (Div. 2)Document4 pagesA. Two Subsequences: Codeforces Round #751 (Div. 2)Trần Nhật KhánhNo ratings yet
- Suretyship and Guaranty CasesDocument47 pagesSuretyship and Guaranty CasesFatima SladjannaNo ratings yet
- 3 Affect Regulation, Mentalization, and The Development of The Self (Peter Fonagy, Gyorgy Gergely, Elliot L. Jurist Etc.) - 211-259Document49 pages3 Affect Regulation, Mentalization, and The Development of The Self (Peter Fonagy, Gyorgy Gergely, Elliot L. Jurist Etc.) - 211-259Sergio Andres Rico AvendanoNo ratings yet
- Green Building ToolsDocument107 pagesGreen Building ToolsVholts Villa VitugNo ratings yet
- Sample Exegesis Paper For IBSDocument12 pagesSample Exegesis Paper For IBScbran250100% (1)
- Prabhupada Sannyasa BookDocument221 pagesPrabhupada Sannyasa BookOriol Borràs FerréNo ratings yet
- Article 405: The Penal-Code With AmendmentsDocument2 pagesArticle 405: The Penal-Code With Amendmentsropal salamNo ratings yet
- First Exam Fifa CorruptionDocument3 pagesFirst Exam Fifa Corruptionarmy kaNo ratings yet
- Eating Disorders Reading AssignmentDocument2 pagesEating Disorders Reading AssignmentNasratullah sahebzadaNo ratings yet
- Standard Procedure For Acoustic Emission TestDocument10 pagesStandard Procedure For Acoustic Emission TestprendoascroccoNo ratings yet
- PhilosophyDocument1 pagePhilosophyElena Luneta-BibeNo ratings yet
- Fire Safety Al RaiDocument4 pagesFire Safety Al RairyecunarNo ratings yet
- Duty of Care Breaches in ASIC v Vines 2006Document8 pagesDuty of Care Breaches in ASIC v Vines 2006sawani mendisNo ratings yet
- Whats New SAP HANA Platform Release Notes enDocument66 pagesWhats New SAP HANA Platform Release Notes enJohn Derteano0% (1)
- Olberding - Other People DieDocument19 pagesOlberding - Other People DiefelmmandoNo ratings yet