You might also like
- Xu, G-H. Et Al 2014 Pteronisculus Nielseni Sp. Nov., A New Stem-Actinopteran Fish From The Middle Triassic of Luoping, Yunnan Province, ChinaDocument17 pagesXu, G-H. Et Al 2014 Pteronisculus Nielseni Sp. Nov., A New Stem-Actinopteran Fish From The Middle Triassic of Luoping, Yunnan Province, ChinaMyrthe de GoeijNo ratings yet
- A New Genus and Species of Scorpionfly (Mecoptera) From Baltic Amber, With An Unusually Developed Postnotal OrganDocument6 pagesA New Genus and Species of Scorpionfly (Mecoptera) From Baltic Amber, With An Unusually Developed Postnotal Organagnieszka.soszynska.majNo ratings yet
- Yang HR 2021 Cretaceous Enigmatic Insect Magicivena SPPDocument11 pagesYang HR 2021 Cretaceous Enigmatic Insect Magicivena SPP杨楠No ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- A New Cretaceous Thyreophoran From Patagonia Supports A South American Lineage of Armoured DinosaursDocument12 pagesA New Cretaceous Thyreophoran From Patagonia Supports A South American Lineage of Armoured DinosaursDino ManiacNo ratings yet
- Letter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaDocument4 pagesLetter: Diverse Transitional Giant Fleas From The Mesozoic Era of ChinaThiago MarinhoNo ratings yet
- A Bizarre Jurassic Maniraptoran TheropodDocument14 pagesA Bizarre Jurassic Maniraptoran TheropodKevin Diego Hernandez MenaNo ratings yet
- A - Gigantic - Feathered - Dinosaur - From - The - Lower - Cretacious Yutyranus PaperDocument5 pagesA - Gigantic - Feathered - Dinosaur - From - The - Lower - Cretacious Yutyranus PapernomadNo ratings yet
- 25 Nature07671 TitanoboaDocument5 pages25 Nature07671 TitanoboaLuisa Fernanda MartinezNo ratings yet
- Po/arornis,: The Morphology and Systematics of A Cretaceous Loon (Aves: Gaviidae) From AntarcticaDocument34 pagesPo/arornis,: The Morphology and Systematics of A Cretaceous Loon (Aves: Gaviidae) From AntarcticaAmelia KumalawatiNo ratings yet
- A New Diapsid From The Middle Triassic of Southern China (Pectodens)Document7 pagesA New Diapsid From The Middle Triassic of Southern China (Pectodens)nooneimportant1986No ratings yet
- The First Dinosaurs From The Early Cretaceous Hami Pterosaur Fauna, ChinaDocument17 pagesThe First Dinosaurs From The Early Cretaceous Hami Pterosaur Fauna, ChinaSebastian IbronNo ratings yet
- PNAS CaiporaDocument5 pagesPNAS CaiporaWalter HartwigNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- Lab 1Document17 pagesLab 1NoyaNo ratings yet
- The Anatomical Record - 2024 - Desojo - Cranial Osteology and Paleoneurology of Tarjadia Ruthae An ErpetosuchidDocument35 pagesThe Anatomical Record - 2024 - Desojo - Cranial Osteology and Paleoneurology of Tarjadia Ruthae An ErpetosuchidCrocodilo VasconcelosNo ratings yet
- Id367 PDFDocument26 pagesId367 PDFannisa nurNo ratings yet
- The Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyDocument20 pagesThe Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyAquaponics OaxacaNo ratings yet
- A New Upper Pleistocene Tapir From ArgenDocument8 pagesA New Upper Pleistocene Tapir From ArgenSebastian QuaroneNo ratings yet
- 2011 Tsuihiji Et Al. Tarbosaurus Juvenile Skull PROOFDocument21 pages2011 Tsuihiji Et Al. Tarbosaurus Juvenile Skull PROOFKanekoNo ratings yet
- Agrostaleyrodes Arcanus A New Genus and SP of MB From TaiwanDocument10 pagesAgrostaleyrodes Arcanus A New Genus and SP of MB From TaiwanJulian LeonardoNo ratings yet
- A Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsDocument19 pagesA Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsHector LealNo ratings yet
- Pleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoFrom EverandPleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoNo ratings yet
- ANATOMIDocument8 pagesANATOMIMaulida Suci AyomiNo ratings yet
- Belokobylskij 2008 Ryukyuspathius Gen NovDocument7 pagesBelokobylskij 2008 Ryukyuspathius Gen NovDávid RédeiNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- Letter: An Enigmatic Plant-Eating Theropod From The Late Jurassic Period of ChileDocument10 pagesLetter: An Enigmatic Plant-Eating Theropod From The Late Jurassic Period of ChilegasfNo ratings yet
- 2003 App48-261 ArchaeoceratopsDocument12 pages2003 App48-261 ArchaeoceratopsKanekoNo ratings yet
- Phylogeny and Biogeography of Asian Pit VipersDocument32 pagesPhylogeny and Biogeography of Asian Pit VipersHaydee ParasNo ratings yet
- The Earliest Herbivorous Marine ReptileDocument6 pagesThe Earliest Herbivorous Marine Reptileyou knowNo ratings yet
- Yan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaDocument12 pagesYan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaCarlos A. JulioNo ratings yet
- 2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDocument16 pages2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDredeltoNo ratings yet
- Avian Evolution: The Fossil Record of Birds and its Paleobiological SignificanceFrom EverandAvian Evolution: The Fossil Record of Birds and its Paleobiological SignificanceNo ratings yet
- Main Menu: Create Account Log inDocument48 pagesMain Menu: Create Account Log inKESATRIA JAKARTANo ratings yet
- A New Species of Polychaete Worm, (Polychaeta: Nereididae) From SingaporeDocument7 pagesA New Species of Polychaete Worm, (Polychaeta: Nereididae) From SingaporeN.R SquireNo ratings yet
- The Relict Cerithiid Prosobranch, (Crosse) : Gourmya GourmyiDocument3 pagesThe Relict Cerithiid Prosobranch, (Crosse) : Gourmya GourmyiLukewarmNo ratings yet
- Condorraptor CurrumiliDocument24 pagesCondorraptor CurrumiliBibbidyNo ratings yet
- A New Basal Coelurosaur Dinosauria TheroDocument16 pagesA New Basal Coelurosaur Dinosauria TheronomadNo ratings yet
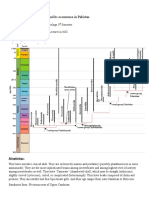
- Cephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanDocument2 pagesCephalopods: Introduction:: Paleontology: Assignment#5 Introduction To Cephalopods and Its Occurrence in PakistanEEsha AhmedNo ratings yet
- Mayr Et Al-2016-PalaeontologyDocument11 pagesMayr Et Al-2016-PalaeontologyJorge BarNo ratings yet
- Beetles Insecta ColeopteraDocument14 pagesBeetles Insecta Coleopteradipeshsingh563No ratings yet
- 2 PDFDocument8 pages2 PDFanon_243369186No ratings yet
- Acta Palaeontologica Romaniae V.4Document16 pagesActa Palaeontologica Romaniae V.4Mada TeodorNo ratings yet
- 02 Chapter 1Document18 pages02 Chapter 1Fabio FotografNo ratings yet
- Theropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRDocument3 pagesTheropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRagomezgallardoNo ratings yet
- A Pictorial Atlas of Fossil Remains: Illustrations from Parkinson's "Organic remains of a former world," and Artis's "Antediluvian phytology"From EverandA Pictorial Atlas of Fossil Remains: Illustrations from Parkinson's "Organic remains of a former world," and Artis's "Antediluvian phytology"No ratings yet
- Camuflage en Insectos - PNASDocument17 pagesCamuflage en Insectos - PNASfany_682403890No ratings yet
- Chordate Evolution and The Three-Phylum System: ReviewDocument10 pagesChordate Evolution and The Three-Phylum System: Reviewnatalia_zimicz1412No ratings yet
- A New ?lamellipedian Arthropod From The Early Cambrian Sirius Passet Fauna of North GreenlandDocument7 pagesA New ?lamellipedian Arthropod From The Early Cambrian Sirius Passet Fauna of North GreenlandSukma MaulidaNo ratings yet
- Evolution of The Sirenia: An OutlineDocument10 pagesEvolution of The Sirenia: An Outlinelahsivlahsiv684No ratings yet
- Functional Anatomy 2019Document28 pagesFunctional Anatomy 2019Ana GiraldoNo ratings yet
- Elpistostege and The Origin of The Vertebrate: ArticleDocument13 pagesElpistostege and The Origin of The Vertebrate: ArticleSevim AkçağlarNo ratings yet
- Product Task OriginalDocument21 pagesProduct Task OriginalHeidi BrionesNo ratings yet
- Spiders of the World: A Natural HistoryFrom EverandSpiders of the World: A Natural HistoryRating: 3 out of 5 stars3/5 (2)
- D1. Neurologic Examination PDFDocument33 pagesD1. Neurologic Examination PDFRamona-Stela CzakoNo ratings yet
- Body Part Reflective EssayDocument4 pagesBody Part Reflective EssayAshley JohnsNo ratings yet
- Full Course Material Can Be Downloaded FromDocument21 pagesFull Course Material Can Be Downloaded FromMuthu JSSCPTNo ratings yet
- Physical Therapy in Sport: Mark J. Stokes, Jeremy Witchalls, Gordon Waddington, Roger AdamsDocument10 pagesPhysical Therapy in Sport: Mark J. Stokes, Jeremy Witchalls, Gordon Waddington, Roger AdamsSaiful AnamNo ratings yet
- Kim - 2015 - Foot Pressure Analysis of Adults With Flat and Normal Feet at Different Gait Speeds On An Ascending SlopeDocument3 pagesKim - 2015 - Foot Pressure Analysis of Adults With Flat and Normal Feet at Different Gait Speeds On An Ascending SlopeAwatef FituriNo ratings yet
- Trigen Humeral 01363v10114 UsDocument44 pagesTrigen Humeral 01363v10114 Useduardo1314No ratings yet
- BBB Month 31 Workout PlanDocument11 pagesBBB Month 31 Workout PlanluniblueNo ratings yet
- IASTM - jOURNALDocument12 pagesIASTM - jOURNALviorelNo ratings yet
- Cubital Fossa PDFDocument18 pagesCubital Fossa PDFKyle TongolNo ratings yet
- Core Training: Course Pack For Pe 122: Fitness ExerciseDocument5 pagesCore Training: Course Pack For Pe 122: Fitness ExerciseJohn Rey Dela PenaNo ratings yet
- Distosia BahuDocument48 pagesDistosia BahuFardimayantiNo ratings yet
- Skin TractionDocument14 pagesSkin TractionAnis IzzatiNo ratings yet
- Congenital Muscular TorticollisDocument19 pagesCongenital Muscular TorticollisRyana AmazonaNo ratings yet
- Juvenile Osteology ReportDocument2 pagesJuvenile Osteology ReportKamren MoradiNo ratings yet
- Therapeutic Exercise Foundations and Techniques 5th Ed Part3Document88 pagesTherapeutic Exercise Foundations and Techniques 5th Ed Part3Alice GiffordNo ratings yet
- PhysioEx Exercise 2 Activity 7Document5 pagesPhysioEx Exercise 2 Activity 7Pavel MilenkovskiNo ratings yet
- Misfit Athletics Hspu Weakness Template 2.0Document8 pagesMisfit Athletics Hspu Weakness Template 2.0Anonymous BEEeTPqiNo ratings yet
- Skull Lessn PlanDocument10 pagesSkull Lessn Plananon_132081100% (1)
- CPC Exam QuestionsDocument3 pagesCPC Exam QuestionshevinpatelNo ratings yet
- Muscle Activation During Exercises To Improve TrunkDocument9 pagesMuscle Activation During Exercises To Improve TrunkYoh ChenNo ratings yet
- Course TA1 Jeffrey Okeson Management of TMDDocument6 pagesCourse TA1 Jeffrey Okeson Management of TMDheidyNo ratings yet
- Tortikolis 2 PDFDocument4 pagesTortikolis 2 PDFwie_wie_wieNo ratings yet
- Clinical Examination of A Case of ParaplegiaDocument27 pagesClinical Examination of A Case of Paraplegiaswathi bs0% (2)
- The Scope of OrthopaedicsDocument12 pagesThe Scope of OrthopaedicsTumbal BroNo ratings yet
- Cast CareDocument4 pagesCast CarernrmmanphdNo ratings yet
- Pathophysiology FracturesDocument2 pagesPathophysiology FracturesSewyel Garburi71% (7)
- Low Back Pain - Differential Diagnosis, Prognosis and TreatmentDocument4 pagesLow Back Pain - Differential Diagnosis, Prognosis and TreatmentFifi TandionNo ratings yet
- Articles by Dr. Nata ParnesDocument19 pagesArticles by Dr. Nata ParnesWatertown Daily TimesNo ratings yet
- GRAVES EDITED IE 1 FinalDocument11 pagesGRAVES EDITED IE 1 FinalasdNo ratings yet
- Choose From The Word Bank Below To Identify This Silly Skeleton's BonesDocument2 pagesChoose From The Word Bank Below To Identify This Silly Skeleton's Bonesbjy525No ratings yet