You might also like
- The Acupuncture Prescription:: A Holistic Approach to HealthFrom EverandThe Acupuncture Prescription:: A Holistic Approach to HealthNo ratings yet
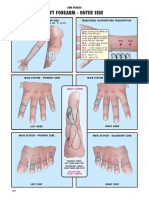
- Hand Shao Yin Vs Hand Jue Yin Wang Hong MinDocument3 pagesHand Shao Yin Vs Hand Jue Yin Wang Hong MinIvonne Flores FernándezNo ratings yet
- Injuries in KarateDocument10 pagesInjuries in Karatemaina_luis6031No ratings yet
- Qi Gong 1Document1 pageQi Gong 1Anass LyamaniNo ratings yet
- Medicine The American Journal of SportsDocument5 pagesMedicine The American Journal of SportsTaregh KaramiNo ratings yet
- TCM Diabetes Treatment ProtocolDocument1 pageTCM Diabetes Treatment ProtocolSreekanth KrishnamurthyNo ratings yet
- Neurophysiology Reflexes) & Osteopathic Manipulative TheoryDocument18 pagesNeurophysiology Reflexes) & Osteopathic Manipulative TheoryCleber TuicciNo ratings yet
- Discover The Beauty of Traditional Chinese MedicineDocument46 pagesDiscover The Beauty of Traditional Chinese MedicinePhuongNo ratings yet
- Ten Important Symptoms in Chinese MedicineDocument100 pagesTen Important Symptoms in Chinese MedicineSuhana hidayatNo ratings yet
- Treatment of Acute Syndromes With Acu and MoxaDocument108 pagesTreatment of Acute Syndromes With Acu and MoxachoylifutsoccerNo ratings yet
- Hering-Breuer ReflexDocument2 pagesHering-Breuer ReflexNTA UGC-NETNo ratings yet
- A Study of The Hering-Breuer ReflexDocument19 pagesA Study of The Hering-Breuer ReflexDenise MathreNo ratings yet
- Kappō 活法: George Arrington Home PageDocument1 pageKappō 活法: George Arrington Home PagehhshshNo ratings yet
- MokusoDocument4 pagesMokusorappa84No ratings yet
- 9780443049767Document15 pages9780443049767Angela PagliusoNo ratings yet
- Therapy ManualDocument129 pagesTherapy ManualValentin HuciuNo ratings yet
- Lachman David Gene ReportDocument108 pagesLachman David Gene ReportDavid LachmanNo ratings yet
- Acupuncture For Children With Autism Spectrum DisorderDocument3 pagesAcupuncture For Children With Autism Spectrum DisorderPaul NguyenNo ratings yet
- Feeding The Hungry GhostsDocument4 pagesFeeding The Hungry GhostsSean BrownNo ratings yet
- Australian Journal Of: Acupuncture and Chinese MedicineDocument60 pagesAustralian Journal Of: Acupuncture and Chinese Medicinepeter911x2134No ratings yet
- Integration of Acupuncture As A Rehabilitation Model Applied in The Integral University Clinic of The State University of The Ecatepec ValleyDocument7 pagesIntegration of Acupuncture As A Rehabilitation Model Applied in The Integral University Clinic of The State University of The Ecatepec ValleyIJAERS JOURNALNo ratings yet
- 10 Leccion 715055 - 635278476265828061Document10 pages10 Leccion 715055 - 635278476265828061angel 1958No ratings yet
- Quiz 1 Study Questions TCM DiagnosticsDocument19 pagesQuiz 1 Study Questions TCM DiagnosticsCherryNo ratings yet
- Chiropractic Technique Principles and Procedures 3Document22 pagesChiropractic Technique Principles and Procedures 3Colors of LifeNo ratings yet
- Acute Spinal Cord Injury Tetraplegia and Paraplegia in Small Animals 2014 Veterinary Clinics of North America Small Animal PracticeDocument26 pagesAcute Spinal Cord Injury Tetraplegia and Paraplegia in Small Animals 2014 Veterinary Clinics of North America Small Animal PracticeJaneKarllaNo ratings yet
- Su Jok and Moxa, A Self-Treatment Manual (PDFDrive) ExportDocument7 pagesSu Jok and Moxa, A Self-Treatment Manual (PDFDrive) ExportJorin JorinNo ratings yet
- Surgical Anatomy of The Face: Implications For Modern Face-Lift TechniquesDocument11 pagesSurgical Anatomy of The Face: Implications For Modern Face-Lift TechniquesDrhisham AttiaNo ratings yet
- DTM Trigger-Point-Referred-Pain-Guide-Interactive PDFDocument439 pagesDTM Trigger-Point-Referred-Pain-Guide-Interactive PDFleokeoNo ratings yet
- Mapping The Body: Space, Time and Gender: Catherine Despeux - 9789004366183 Via Free AccessDocument18 pagesMapping The Body: Space, Time and Gender: Catherine Despeux - 9789004366183 Via Free AccesscarmelaNo ratings yet
- New Zealand School of Acupuncture and TraditionalDocument12 pagesNew Zealand School of Acupuncture and Traditionalfann80100% (1)
- The History of Chinese Medicine in The People S Republic of China and Its GlobalizationDocument21 pagesThe History of Chinese Medicine in The People S Republic of China and Its GlobalizationJumboNo ratings yet
- A Clinician's Guide To Using Granule ExtractsDocument252 pagesA Clinician's Guide To Using Granule ExtractsSalsa Nabila AwliaNo ratings yet
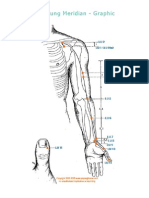
- (LU) Lung Meridian - GraphicDocument16 pages(LU) Lung Meridian - GraphicNeacsu TudorelNo ratings yet
- Chen Bing PDFDocument26 pagesChen Bing PDFHankNo ratings yet
- Xin Yan Et Al - External Qi of Yan Xin Qigong Differentially Regulates The Akt and Extracellular Signal-Regulated Kinase Pathways and Is Cytotoxic To Cancer Cells But Not To Normal CellsDocument12 pagesXin Yan Et Al - External Qi of Yan Xin Qigong Differentially Regulates The Akt and Extracellular Signal-Regulated Kinase Pathways and Is Cytotoxic To Cancer Cells But Not To Normal CellsSonyRed100% (1)
- Pulse Diagnosis: Martha Lucas 2nd Ed.Document143 pagesPulse Diagnosis: Martha Lucas 2nd Ed.Vita MishtiiNo ratings yet
- Treatment of Hyperthyroidism: Western Medicine vs. Traditional Chinese MedicineDocument6 pagesTreatment of Hyperthyroidism: Western Medicine vs. Traditional Chinese MedicineShahul HameedNo ratings yet
- 2009 The Neuroanatomic Basis of The Acupuncture Principal MeridiansDocument21 pages2009 The Neuroanatomic Basis of The Acupuncture Principal MeridiansAna Maria MartinsNo ratings yet
- To Understand Moxibustion From The Biological Effect of Local Thermal StimulationDocument8 pagesTo Understand Moxibustion From The Biological Effect of Local Thermal StimulationMiki ShimizuNo ratings yet
- Home About Us Order Info FAQ Distributors Suppliers Feedback Contact Us View CartDocument5 pagesHome About Us Order Info FAQ Distributors Suppliers Feedback Contact Us View CartdpscribdzoneNo ratings yet
- (Apostila) ACUPUNTURA. Leituras Científicas 1 PDFDocument66 pages(Apostila) ACUPUNTURA. Leituras Científicas 1 PDFEvelyn GolinNo ratings yet
- Brennan Translation of Xu Yusheng's 18 Luohan ExercisesDocument6 pagesBrennan Translation of Xu Yusheng's 18 Luohan ExercisesRoberto BaptistaNo ratings yet
- Chinese Medical Qigong GuideDocument1 pageChinese Medical Qigong GuideLaura FerrariNo ratings yet
- Diagnostic Protocol ModificatDocument83 pagesDiagnostic Protocol Modificatuda20130% (1)
- Auriculotherapy Manual: Chinese and Western Systems of Ear AcupunctureDocument2 pagesAuriculotherapy Manual: Chinese and Western Systems of Ear AcupunctureYh KooNo ratings yet
- Ryodoraku History and ResearchDocument6 pagesRyodoraku History and ResearchKate RyncaNo ratings yet
- (OHNS) Dizziness and Vertigo-Dr. Yang (JM Sales)Document8 pages(OHNS) Dizziness and Vertigo-Dr. Yang (JM Sales)Noreen Hannah GabrielNo ratings yet
- Dr. Yossi Maryanti, M.biomed, Sp. S - Neck and Back PainDocument50 pagesDr. Yossi Maryanti, M.biomed, Sp. S - Neck and Back PainFreade AkbarNo ratings yet
- Taiji and Taiji QuanDocument9 pagesTaiji and Taiji Quanapi-311414449No ratings yet
- Acupoint Combination Methods LectureDocument27 pagesAcupoint Combination Methods LectureUdhayNo ratings yet
- Acupuncture: Ancient Treatment For Modern IllnessesDocument31 pagesAcupuncture: Ancient Treatment For Modern IllnessesJessica PaulNo ratings yet
- Meridians linked to infrared changesDocument5 pagesMeridians linked to infrared changesAhmed HusseinNo ratings yet
- Pulse Diagnosis Device enDocument7 pagesPulse Diagnosis Device enYj XuNo ratings yet
- CatalogueDocument292 pagesCatalogueTiit RaagNo ratings yet
- ATCM JOURNAL September 2014 - 21 PDFDocument60 pagesATCM JOURNAL September 2014 - 21 PDFIvonne Flores FernándezNo ratings yet
- The Journal of The Rolf Institute March 2017Document56 pagesThe Journal of The Rolf Institute March 2017Taune AntonioNo ratings yet
- Zen ShiatsuDocument8 pagesZen ShiatsuIoan-ovidiu CordisNo ratings yet
- HHS Public Access: Nerve Stimulation: Immunomodulation and Control of InflammationDocument28 pagesHHS Public Access: Nerve Stimulation: Immunomodulation and Control of InflammationAdán LópezNo ratings yet
- 1 s2.0 S1548559505000765 MainDocument10 pages1 s2.0 S1548559505000765 MainosmarcruzNo ratings yet
- Acupuncure and ImmunomodulationDocument14 pagesAcupuncure and Immunomodulationcristinacaetano9355No ratings yet
- Lucid Dreaming - Experience and Control Your DreamsDocument2 pagesLucid Dreaming - Experience and Control Your DreamsMistermickeyNo ratings yet
- BFCRS With Manual LinksDocument1 pageBFCRS With Manual Linkskirti thakurNo ratings yet
- Adhd Referral CriteriaDocument2 pagesAdhd Referral CriteriaPutryy FitrilyaaNo ratings yet
- Theories of Intelligence SeminarDocument32 pagesTheories of Intelligence SeminarJonathan L SabaNo ratings yet
- FS1 Semi FinalsDocument16 pagesFS1 Semi FinalsAsh LafuenteNo ratings yet
- Theories of Personality NOTESDocument17 pagesTheories of Personality NOTEStaylor alison swiftNo ratings yet
- Perception and LearningDocument82 pagesPerception and Learningnaveen nani100% (1)
- Sharot Korn Dolan How Unrealistic Optimism Maintained in The Face of Ukmss-36575 PDFDocument15 pagesSharot Korn Dolan How Unrealistic Optimism Maintained in The Face of Ukmss-36575 PDFAnonymous pb0lJ4n5jNo ratings yet
- Jurisprudence Direct Exam of Expert WitnessDocument9 pagesJurisprudence Direct Exam of Expert WitnessxquisiteDNo ratings yet
- Difference Between Training and DevelopmentDocument2 pagesDifference Between Training and DevelopmentMoe NyiNo ratings yet
- The Milan Approach - Roots and EvolutionDocument4 pagesThe Milan Approach - Roots and EvolutionVivir Sin ViolenciaNo ratings yet
- Assessing Mental Status and Well-BeingDocument7 pagesAssessing Mental Status and Well-BeingAnnalisa TellesNo ratings yet
- Reading As A Physiological ProcessDocument2 pagesReading As A Physiological ProcessRho Vince Caño Malagueño89% (9)
- YOLO V3 ML ProjectDocument15 pagesYOLO V3 ML ProjectAnnie ShuklaNo ratings yet
- ASD and AnxietyDocument87 pagesASD and AnxietyH.JulcsiNo ratings yet
- Mam Gene ReportDocument13 pagesMam Gene ReportOhmel VillasisNo ratings yet
- Chapter 3Document7 pagesChapter 3Generose GamayNo ratings yet
- The BACKPROPAGATION Algorithm: KD KDDocument10 pagesThe BACKPROPAGATION Algorithm: KD KDSwathi ReddyNo ratings yet
- Ultimate Brain CDs ReviewDocument6 pagesUltimate Brain CDs ReviewUjjwal Rastogi100% (1)
- Brain Computer Interface Technical Seminar ReportDocument11 pagesBrain Computer Interface Technical Seminar ReportA.srikanth100% (1)
- Social isolation nursing diagnosis schizophreniaDocument3 pagesSocial isolation nursing diagnosis schizophreniaChuck Smith100% (11)
- Acceptance and Commitment Therapy Unified Model Behavior Change PDFDocument27 pagesAcceptance and Commitment Therapy Unified Model Behavior Change PDFRENE TOMATEO CONTRERASNo ratings yet
- Dyslexia - Disability or Gift?: Savitri Dias University of Colombo Sri LankaDocument25 pagesDyslexia - Disability or Gift?: Savitri Dias University of Colombo Sri Lankadias_savitriNo ratings yet
- Ineco Social BrissDocument8 pagesIneco Social BrissPau RochaNo ratings yet
- Module 4 - T EnglishDocument11 pagesModule 4 - T EnglishGlaiza OrpianoNo ratings yet
- Motivation: William G. HuittDocument14 pagesMotivation: William G. Huittamritam yadavNo ratings yet
- FlashtrainingfiftheditionDocument14 pagesFlashtrainingfiftheditionEvening PrimeNo ratings yet
- LOCUS scoring sheet analysisDocument2 pagesLOCUS scoring sheet analysistakemebrickNo ratings yet
- What Do Children Learn About Prosocial Behavior From The Media?Document6 pagesWhat Do Children Learn About Prosocial Behavior From The Media?Heather Edey WilliamsNo ratings yet
- Peterson 2005Document17 pagesPeterson 2005Пламен МинчевNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (14)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (327)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Daniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisFrom EverandDaniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisRating: 3.5 out of 5 stars3.5/5 (130)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisFrom EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisRating: 5 out of 5 stars5/5 (3)