16 SECTIUNEAAIEN + Fig cer gmeleee
transmembranare sunt molecule
este mult: mai
Deoarece_protenee
aia rf ml
cr in plus, de fixarea lor pe citoschelet, chiar
aaa eke episeze In plan erate
ar sant mult mal pide ma pin dre:
sro sor det ifrunen Protenee motor, cate sunt
sociale ct ctoscheletul cituplasiiati (diseutate mai jos), par
aS Sacha pocion Sessmmnbonee, cghrate
fiznal membrane precum Juciile barca cae plese
fn siruri, In cele din urma, proteinele pot difuza numai in
ploul Bsa precum fosoipidele Eien pot ravers
See snes pera glans damier ecco
Ghoplenelce bite one! potsine tansmernbranare
peste nucleul hidrofob al bistratului este foarte dificil de sur-
ont es trae topolo el en
Semodifia dea lng dare! sale
ve membranare nit
FUNCTIA PROTEINELOR MEMRRANARF.
Proteinele membranare integrale pot servi ca receptori
‘Yoate comunicarile dintre o celula si mediul sau inconjurator
trebuie si implice, sau cal putin 28 troverseze membrana
plasmatica. In scopul acestei discuti, definim comunicarea
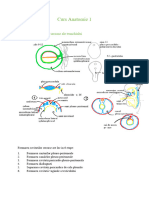
A REGEPTORII DE LEGARE LIGAND
Domeniu de legare igand
7 segmente
srensmembronere
if
Tigwett veie meneranare eg cae tans or
post fun heen, un actor dees noocuarera
Iain ete © molec in dso cre cages cha Se
ito, un
In sens ha bul oricirui cemnal intre celula g
ceva ce o inconjoara. Cu exceptia moleculelor semnal soly.
bile in lipice, cum sunt liormonii steroizi, in event, toate
Fanctiile de comunicare deservite de membrana plasmaticg
se realizeari prin intermediul protemelor membranare,
Dintr-v perspectiva de inginerie, proteinele membranare
sunt situate perfect pentru a transmite semnale, deoarece ele
formeaza o legitura continua, unica, intre doua comparti.
inet ve sunt separate de membran
-ceptorii de legare ligand cuprind grupnl proteinelor
transmembranare, care pot ilustra, probabil, cel mai clar
conceptul de scmnalizare transmembranari (Fig. 2-7).
Pentru ca hormonii solubili in apa, cum este epinetrina, si
influenfeze comportamentul celular, prezenfa lor in com-
partimentul fluidului extracelular (ECE) trebuie si fe facut
Cunoscuta diferitelor mecanisme intracelulare, ale caror
comportamente le moduleaza. Interactiunea unui hormon
cu portiunea extracelulard a receptorului hormonal. care for-
rmeazi un situs de legare cu mare afinitate, produce sel
iri canformatianale in proteina receptor, care ce eetind
domeniul intramembranar la domeniul intracelular al
ji, Ca urmare, domeniul intracelular, fie devine
activ din punct de vedere enzimatic, fie poate interactiona cu
proteinclecitoplarmatice care cunt implicate in generarca aja
‘umitlor mesageri secun2i. Fiecare mecanism completeaza
transmisia semnalului hormonului de-a lungul membranci.
Dispozitia transmembranara a receptorulut hormonal for-
‘meszi ectfel, un mediu de comunicare continus unie, care
este capabil de a transmute, prin proprile sale modificiri
MOLECULA DE ADEZIUNE LA MATHICEA CELULAHA,
(NTEGRINA)
7 Domeniu de legare matrice
Domenie
citoplasmatice sunt
logate de proteine
intracelulare.
loin terior spre nterionl cell A, Ligand
codorizant sau un alt mediator ‘
ices extracelulars, ees
a
Scanned by CamScanner
urale, intormatii din mediul extracelular interiorului
celular. Procesul de transductie al semnalului transmembra-
nar este discutat in Capitolul 3,
Proteinele membranare integrale pot servi ca molecule de
adeziune
Celulele pot, de asemenea, exploata proteinele membranare
integrale, utilizinducle ca molecule de adeziune care for-
smeazi contact fiice cu matricea extracelular inconjuritoare
(ex. molecule de adeziune celuli-matrice), sau cu vecinii lor
celular (ex, molecule de adeziune eelulé-celli), Acesteataga-
mente pot fi extrem de importante in reglarea formei,cresterit,
i diferentieri celulelor. Natura gi amploarea aceztor ataga-
mente trebuie si fie comunicate in interiorul celulei, astfel
{ncit celula si se poati adapta adecvat la constringerile fizce
i indicile care sunt furnizate de mediul imediat inconjurator.
‘Numeroase clase de proteine transmembranare sunt implicate
fn aceste procese de comunicare. Integrinele sunt exemple de
receptori matriceal sau molecule deaderiune celuli-matrice.
Ele cuprind o familie mare de proteine transmembranare, care
leagi celulele la componente ale matricei extracclulare (ex.
hibronectina, laminina) 1a nivelul placilor de adeauune (vet
Big. 2-78), Aceste legituri produc modificiri conformationale
{in moleculele de integrine, care sunt transmise cozilorlor cito-
plasmatice. Acestecozi, la rindul lor, comunici evenimentele
de legare diverselor molecule structurale gi de semnelizare,
care participa la formularea unui raspuns al celles la mediul
8 fiaie inconjuritor.
Spre deosebire de receptorii matriceali, cate atageazd
celulele 1a matrices extracelular’, mai multe superfamilii
enorme de molecule de adeziune celuli-celula atajeazi
celulele intre ele, Aceste molecule de adeziune celula-ce-
lula includ moleculele de adeziune celulari Ca”* dependente
(Caderinele) si moleculele de adeziune celulara neuronale
Ca independente (N-CAMs). Caderinele sunt glicaprote-
ine (adica, proteine cu glucide atagate) cu un segment intra-
membrana si un domeniu larg extracelular care leaga C2"
N-CAMs, pe de alti parte, in general, sunt membri ai super-
familiei de imunoglobuline. Cele doud clase de molecule de
‘adeziune celuli.celuls. media74 tipuri similare de semnale
‘ransmembranare, care ajuté la organizarea citoplasmei si
expresia genelor de control, ca rispuns la contactele inter-
celulare. Unele molecule de adeziune celula-celula apargin
clasei GPI-linkate de proteine membranare (vezi pag, 13).
Aceste polipeptide nus au coadi tranemembranari si cito-
plasmaticd. Interactiunile mediate de aceasta clasi unicé de
molecule de adeziune pot fi comunicate in interiorul celulet,
prin asocieri laterale cu alte proteine membranare.
Moleculele de adeziune orchestreaz§ procesele foarte
diverse, precum migrarea directionat® a eelilelor imunitare
si ghidarea axonilor in sistemu! nervos in curs de dezvoltare.
Pierderea adeziunilor celuli-celul si celuli-matrice este un
semn distinctiv al celulelor tumorale metastatice,
Proteinele membranare integrale pot efectua transportul
{ransmembranar al substanfelorsolubile in apa
Dup& cum am mentionat mai sus, un bistrat fosfolipidie
Pr nu are proprieiile de permeabiltate care suntjin mod
Normal, asociate cu: membranele ;plasmatice: a
CAPTOWUL2 = Owain uncon acehiel 17
animale. Bistraturile fosfolipidice pure, de asemenea, nu
au capacitatea de a transporta substanfe in sens ascendent
(adica impotriva gradientilor electrochimici; pag. 105),
Proteinele transmembranare inzestreari membranele bialo-
give cu aceste capacitili Joni, sialte substane cu membrana
impermeabil, pot traversa bistratul cu ajutorul proteinelor
transmembranare, care servese ca pori, canale, c&riusi i
pompe. Vorit 51 canalele servesc drept conducte care permit
ape, ionilor specifici sau chiar proteinelor foarte mari ?
raBeLUL 5-2 super
mae aga ones IO
fans
rrsportoi de gutamet
1 acitat al nexozelot
ale vansportorilor
je aminoac (eu LCT)
ri de HCO
‘Transport
suite orl
heterodimetiil
serumpaturi 3 cotransport
cotransporter Na/alucord
ovrensportl cupat cu Na" $I #!
“neurowrensmistorior”
ynsportonlor
itil treneportoare @ tra
‘Subunit tranepor ae
hheterodimerci de aminoacizi (6
‘scimatori Na-Ca
‘schimbatori No-H
Ccotransportri Na sérun Diare
Cotransportori onlor metalic controls de
ue
‘Cotranspertri cationici cuplai cu Cl;
Goransportor de sult 9 carboxlat
couple cu Na
Transport faciitat al ureel
Ccouansportl ve uiyopeptide dirt de
i
“Transpartori de monocarboxilat
CCotransportori Na /fostat de tpul | 51
‘eansportor vesicular de Gla
“Tensportot veziculan Ge monoamine
“ranspotor de folate
CCotransoortorit Na fost de tpul i
"anor 6 iin atin gan
Cotenspotai de aro cation
amfoteri organici i
14; HMIT
etectrogeni)
tor No/HICO; electroneut)
aca 2 eatanspor Ne ce Nat)
SGLI1 2,34 § (glue
IS (odid); SMITS 2(ei
‘SMcTi, 2 (enonocarboxta)
SMV F (iting)
CHT (colind)
Part (aminoacid vuplet cu Na")
GAT 2, 3: GBT! (GABA cuplat cu No
Sre*=(aminoacia cupali cu Na” $i CI)
NET (transportori norepineftinel)
SERT (serotonind)
DAT (dopamin8)
Lary 2; y"LATL 2
CATI.2.3.4
NOX 2.3
NHE, 2, 34,5, 6,789
NHAI.2
Sperm-NHE
set
own
NRAMPI
NKCC1,2 (eotransportorii Na/KIC!)
NCC (cotrensportorii Na/C!)
Ker, 2, 8 4 (cotransportorii K/CI)
NaDC1, 3 (mono, di i tricarboxli)
NaSt 2 (sult)
UTAL 2,34 5,6; UT-BI 2
opt 2
rm 2 j
MCT, 2, 3,4 (cotransportorul de monocarrrboxilati ct
_ i cuplat cu H"
INCTS otucune cat omnoncalor wematig)
NaPi 7 .
NETH 3.4 : at son
NUT i '
VGLUTL2.3
\VMATI,2 (Schimbstorul H*-amine)
\VACHT (schimbstoru H*-aceticoind)
REC” maa .
Tha,2 td, : *
Pita, 2 .
ATP 2, 3/456 (sari
horn tridien,prostaglandine)
‘OCT 2, 3 (difuziunea facil
, cilitats tic
seh dma crag
Ont Sutarseerteu Nafeation’ organi
Scanned by CamScanner
TABELUL 5-4
FAIL
23 (4)
sca (9)
C28 (53)
$1026 (M)
sczr ©)
102 (3)
s1cz9 (4)
$1030 (10)
scat (2)
stca tt)
$1033 (1)
L034 (3)
3.035 (00)
1036 (4)
Lear (4)
S038 (1)
‘lca (18)
stc40 (1)
sea (3)
stc2(3)
Stes (3)
S104 (6)
Reis
SLC46 (3)
Stour @)
sucte
&
Stes ()
“SUCBTTQyy
SLC52 (9)
1 S16+9(4) _ Transpoitorllegat de FLVOR)
SES
“rensportori Ne 7 acid ascorbic
‘Schimbatorul No*/(Ca"*-K")
CBrBugi mitocondrial
Schimb8torii multifunctional de anion
‘Transpertori de acizi gras
‘Trensportorii Ne /nucleozide
‘Transportorifeciltstori de nucleozide
‘Transportorieflualui de 2n
Importatorul de cupra
‘Tansportorl vesicular inhibitor de aminoaczi
‘ransportorul de acetl-coonzims A
CCotranaportorii Na/fosfat de tpul I
Cotranaportorul zhorul nuclooaidelor
CAPTOLULS + Vansant apelelectoer 193,
NGI, 2,3, 4,
ANCL, 2, 3, 4 (ANTI, 2,3, 4)
DRA (schimbatorul CIHCO3)
endrina (schimb HCO, CI” sau (}
Prestina (gchimba Cl, format, oxalat; un motor in celule ciliate cohleare)
CCFEX (schimbs Cr, HCO, oxalat, frrnat)
FATPL2,3,4,5,0
onty2.3
ENTL2.3,4
ZT 2,3, 4,5, 6,7, 0,9, 10
crm,2
IAAT (schimbitor de Ht cu GABA sau glicind)
ACATNI
NaPi tab, 6
HER
est
uct
PAPSTY, 2
Cotvansportori de aminoacii cuplaticu HY PATI 2.3.4
‘Schimb&torul zahar-fosfat/fosfat
Transportul aminoaciailor neutr dirjat de
Nat (sisterele A si N)
Transportori jonilor metalici
Tronsportorul bazolateral de Fe™
‘Transporter de magnézi MGte-ke
‘Canclele NH /NHi, CO,
Sictomul Lk ol raneportatoior de
‘amino indepedent de No
Transportorl de otns
“rensportorlprosumt de 2aha?
Transportorul de folat
‘Tansportorul multdrog si de'toxine etminate MATES 2
‘Tansportorul hemulul
a poegapveeenteaned
“Tranaportorl fuel de zahar
"-raneprsrl leailor dematé ih
“Traneportorulribofavinei
SPKL2,3,4
SSNATY 2,4 (sistemul A, cotransporta aminoaciz eu Na")
SSNAT3 5 (sistem, cotransperth aminoacii cu Na*laschimb cu H")
ZIP 2,3 4, 66,78, 9,10, 1 12, 13,14 (absorbia Zn™)
Feroporing (FPN!)
Note!
RAG, RNBG, RNCG
EATS oidssnnil Hit
EEG abet pistes
CTL 2.3.4,5
{ince (gene 1 responebil penn he)
vera
MEST”
ie ae ee eee cnaeiner ach
aie gm OE ETE
an TE ri nn at ene SSH rw EIA en
i
vores ete numiarul ue gave unre
cr ttcte fami’ iy este un umd
cid gaba-aminabutine.
“Tus wom ar harden geer st Gnuile SLC (e war SLCAAA wd et in tr
\dontcare agen n cada cee om
Scanned by CamScanner
114 —secTUNERAN lea cele si molcelr
A DIFUZIUNEA SIMPLA 1B DIFUZIUNEA MEDIATA DE
‘CARAUS SAU FACILITATA
4 4
Jac
ran
t
id mek,
Figura 5 Dependenta ratelor de transport de concentraja substente
You might also like
- Cyber ClosetDocument6 pagesCyber Closetalexandra constantinNo ratings yet
- Calitatea ApeiDocument4 pagesCalitatea Apeialexandra constantinNo ratings yet
- Contract de Voluntariat Arges '23.docx NineDocument4 pagesContract de Voluntariat Arges '23.docx Ninealexandra constantinNo ratings yet
- Cursuri AnatomieDocument63 pagesCursuri Anatomiealexandra constantinNo ratings yet
- Mircea EliadeDocument5 pagesMircea Eliadealexandra constantinNo ratings yet
- Probleme Subprogramecls10Document11 pagesProbleme Subprogramecls10alexandra constantinNo ratings yet
- Fizio C4 BoronDocument27 pagesFizio C4 Boronalexandra constantinNo ratings yet