You might also like
- Lesson 5 Structure and Functions of Leaves Student WorksheetDocument3 pagesLesson 5 Structure and Functions of Leaves Student WorksheetEunice Xiiao Wenn100% (2)
- Armour Et Al. - 1997 - Gross and Microscopic Anatomy of The Human Intrinsic Cardiac Nervous SystemDocument10 pagesArmour Et Al. - 1997 - Gross and Microscopic Anatomy of The Human Intrinsic Cardiac Nervous SystemshaddaiboozNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptedwinNo ratings yet
- Anatomy and Physiology of Intrinsic Cardiac Autonomic Nervous SystemDocument5 pagesAnatomy and Physiology of Intrinsic Cardiac Autonomic Nervous SystemSori B. Requena C.No ratings yet
- Nervous System - NSUDocument42 pagesNervous System - NSUGregrodocker100% (1)
- Janes 1986Document11 pagesJanes 1986Luca ClementinoNo ratings yet
- 1995.nerve Cells in The Human Ciliary Muscle. Ultrastructural and Immunocytochemical CharacterizationDocument13 pages1995.nerve Cells in The Human Ciliary Muscle. Ultrastructural and Immunocytochemical CharacterizationReza NaghibiNo ratings yet
- Experimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalDocument13 pagesExperimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalAndré DiasNo ratings yet
- Powerpoint Decompressive CraniectomyDocument33 pagesPowerpoint Decompressive CraniectomySyarif HidayatullahNo ratings yet
- J of Comparative Neurology - 2023 - BizantiDocument22 pagesJ of Comparative Neurology - 2023 - BizantiIgnacio BarbieriNo ratings yet
- Johns2011 - LECTURA RECOMENDADADocument37 pagesJohns2011 - LECTURA RECOMENDADAFRANCISCA ISIDORA ROJASNo ratings yet
- Function Vagal SystemDocument17 pagesFunction Vagal SystemMaura HokamaNo ratings yet
- Chapter Two 10-01-2021Document8 pagesChapter Two 10-01-2021Mustafa AL ShlashNo ratings yet
- Cardiac Neuroanatomy - Imaging Nerves To Define Functional Control PDFDocument40 pagesCardiac Neuroanatomy - Imaging Nerves To Define Functional Control PDFHappy PramandaNo ratings yet
- Physiological Psychology: Physiological Psychology Studies TheDocument18 pagesPhysiological Psychology: Physiological Psychology Studies The31316969No ratings yet
- Nerve Supply of The Human Knee and ItsDocument7 pagesNerve Supply of The Human Knee and ItsErnesto VillavicencioNo ratings yet
- Satellite Article: Horner's Syndrome in HorsesDocument5 pagesSatellite Article: Horner's Syndrome in HorsesJessica AlejandraNo ratings yet
- Zhang Et Al 2023Document21 pagesZhang Et Al 2023Ignacio BarbieriNo ratings yet
- Leavitt 2005Document16 pagesLeavitt 2005qr7bht566gNo ratings yet
- The Enteric Nervous System: A Second BrainDocument14 pagesThe Enteric Nervous System: A Second Brainmatiashernan123No ratings yet
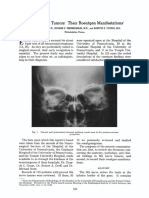
- Eighth Nerve Tumors: Their Roentgen: Manifestations'Document33 pagesEighth Nerve Tumors: Their Roentgen: Manifestations'Elsya PutriNo ratings yet
- Park 2012Document5 pagesPark 2012Arankesh MNo ratings yet
- Histology of Cardiac MuscleDocument13 pagesHistology of Cardiac MuscleAslinNo ratings yet
- Spinal Cord AnatomyDocument34 pagesSpinal Cord AnatomyYana CatherineNo ratings yet
- University of Pennsylvania Neuroscience ResearchDocument25 pagesUniversity of Pennsylvania Neuroscience Researchapi-495461280No ratings yet
- Wong Et Al-2013-The Blood-Brain Barrier An Engineering PerspectiveDocument22 pagesWong Et Al-2013-The Blood-Brain Barrier An Engineering PerspectiveGourav KumarNo ratings yet
- Auer1984 Article TheDistributionOfHypoglycemicBDocument15 pagesAuer1984 Article TheDistributionOfHypoglycemicBJean Pierre Chastre LuzaNo ratings yet
- Autonomic Neuroscience: Basic and ClinicalDocument11 pagesAutonomic Neuroscience: Basic and ClinicalCarlos ZepedaNo ratings yet
- Disorders of The Lower Cranial Nerves: Review ArticleDocument15 pagesDisorders of The Lower Cranial Nerves: Review Articleamelia marhelesNo ratings yet
- Henne Man 1965Document21 pagesHenne Man 1965Brayan BuitragoNo ratings yet
- Cerebral Artery RevisedDocument16 pagesCerebral Artery RevisedbobirfansyahputraNo ratings yet
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDocument34 pagesThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337No ratings yet
- Neuroanatomy Notes 2010Document33 pagesNeuroanatomy Notes 2010Jennifer David InauenNo ratings yet
- Janat00631 0003Document29 pagesJanat00631 0003Luca ClementinoNo ratings yet
- Review Article: Brain Death and Organ DonationDocument6 pagesReview Article: Brain Death and Organ Donationspagete18No ratings yet
- UntitledDocument222 pagesUntitledMyat OoNo ratings yet
- Carr Central ProjectionsDocument13 pagesCarr Central ProjectionsMengda ZhangNo ratings yet
- Husos de SueñoDocument37 pagesHusos de SueñoYair FloresNo ratings yet
- Accepted Manuscript: Seminars in Cell & Developmental BiologyDocument18 pagesAccepted Manuscript: Seminars in Cell & Developmental BiologyJANNETH PRISCILA JACHO TORRESNo ratings yet
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Anatomic Considerations of Superior Laryngeal Nerve During Anterior Cervical Spine ProceduresDocument4 pagesAnatomic Considerations of Superior Laryngeal Nerve During Anterior Cervical Spine ProceduresMark DenhamNo ratings yet
- Brain NeuroanatomyDocument227 pagesBrain NeuroanatomyNkechukwu OgakwuNo ratings yet
- 1-S2.0-S1878875022005460-Main Filum Terminale: A Comprehensive Review With Anatomical, Pathological, and Surgical ConsiderationDocument10 pages1-S2.0-S1878875022005460-Main Filum Terminale: A Comprehensive Review With Anatomical, Pathological, and Surgical ConsiderationYo MeNo ratings yet
- Norvell ARG and Role in Renal InnervationDocument11 pagesNorvell ARG and Role in Renal InnervationGNo ratings yet
- Brainstem and Nuclei Condensed Grayscale SlidesDocument17 pagesBrainstem and Nuclei Condensed Grayscale SlidesMobarobberNo ratings yet
- Introduction To Autonomic PharmacologyDocument9 pagesIntroduction To Autonomic Pharmacologynarindersingh15No ratings yet
- Common Neurologic Disorders: Krista M. GarnerDocument12 pagesCommon Neurologic Disorders: Krista M. GarnerTeda G.No ratings yet
- Physiological Psy LectureDocument125 pagesPhysiological Psy LectureKdrama SweetieNo ratings yet
- Human BrainDocument44 pagesHuman BrainSri vaishnava100% (1)
- Winkler 1889Document3 pagesWinkler 1889chemistpl420No ratings yet
- Cardiac Dysfunction in Neurocritical Care.Document14 pagesCardiac Dysfunction in Neurocritical Care.Angela Moreira ArteagaNo ratings yet
- J Yebeh 2019 106551Document6 pagesJ Yebeh 2019 106551Luisa Fernanda Lopez MoraNo ratings yet
- 1974 Axelrod-NEUROTRANSMITTERSDocument15 pages1974 Axelrod-NEUROTRANSMITTERScortizmenNo ratings yet
- The Nervous System 2Document84 pagesThe Nervous System 2Erika MatiasNo ratings yet
- Jurnal PendidikanDocument8 pagesJurnal PendidikanQothrun Nada FYNo ratings yet
- Parvertebral Blokade of TheDocument11 pagesParvertebral Blokade of TheClínica Veterinaria TODOVETNo ratings yet
- Adverse Neural Tension WorkbookDocument29 pagesAdverse Neural Tension WorkbookCristi MansillaNo ratings yet
- Lennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)Document395 pagesLennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)αργυ αργυ100% (1)
- The Adrenal GlandsDocument34 pagesThe Adrenal Glandsashgee1No ratings yet
- Cellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8From EverandCellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8No ratings yet
- Anevidence-Based Videotapedrunning Biomechanicsanalysis: Richard B. SouzaDocument20 pagesAnevidence-Based Videotapedrunning Biomechanicsanalysis: Richard B. SouzaInfo-qe QualityenergyNo ratings yet
- A Systematic Review of Running-Related Musculoskeletal Injuries in RunnersDocument10 pagesA Systematic Review of Running-Related Musculoskeletal Injuries in RunnersInfo-qe QualityenergyNo ratings yet
- 1 s2.0 S1053811917301015 MainDocument9 pages1 s2.0 S1053811917301015 MainInfo-qe QualityenergyNo ratings yet
- CMJ 129 2373Document8 pagesCMJ 129 2373Info-qe QualityenergyNo ratings yet
- Post-Exercise Changes in BloodDocument9 pagesPost-Exercise Changes in BloodShafie QahNo ratings yet
- Blood Vascular System of Herdmania-part-II-zoologyDocument6 pagesBlood Vascular System of Herdmania-part-II-zoologyPraba KaranNo ratings yet
- Variant Angina & Microvascular AnginaDocument59 pagesVariant Angina & Microvascular AnginaazizhaNo ratings yet
- Basics - ECGpediaDocument6 pagesBasics - ECGpediapakpahantinaNo ratings yet
- Heart - Structure: Left Atrium Right Atrium Left Ventricle Right VentricleDocument3 pagesHeart - Structure: Left Atrium Right Atrium Left Ventricle Right VentricleTharanga HewabuhageNo ratings yet
- Work Book Activity 9 Ana LabDocument7 pagesWork Book Activity 9 Ana LabJazmine YvonneNo ratings yet
- Microbiology: Basic and Clinical Principles: First EditionDocument110 pagesMicrobiology: Basic and Clinical Principles: First EditionDawn DixonNo ratings yet
- Antidiuretic Hormone PDFDocument3 pagesAntidiuretic Hormone PDFMcRo UndiNo ratings yet
- Transportation in Plants and Animals Ncert and Extra QuestionsDocument6 pagesTransportation in Plants and Animals Ncert and Extra QuestionsNaitik MeenaNo ratings yet
- MAGCAR 12 - Cardiopatías CongénitasDocument61 pagesMAGCAR 12 - Cardiopatías CongénitasMovil Maria 2No ratings yet
- Lec.3 PhlebetomyDocument2 pagesLec.3 PhlebetomyQudamah JasemNo ratings yet
- Multiple Assessment Ma - 2, XDocument3 pagesMultiple Assessment Ma - 2, XAarjav JainNo ratings yet
- ThirstDocument30 pagesThirstvalcobacl.681.studNo ratings yet
- BodyBallancer Lipo Brochure PDFDocument2 pagesBodyBallancer Lipo Brochure PDFRoss Gale100% (1)
- Functional Anatomy-RespirationDocument37 pagesFunctional Anatomy-RespirationASIIMWE WINNIE CATHERINENo ratings yet
- TestDocument46 pagesTestmohammed faqihNo ratings yet
- Kami Export - CirculatorySystem12052022Document1 pageKami Export - CirculatorySystem12052022Nyla ScottNo ratings yet
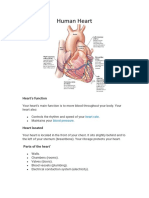
- Human HeartDocument7 pagesHuman HeartAsher Eby VargeeseNo ratings yet
- Wound Healing: Biology Presentation by AbbyDocument8 pagesWound Healing: Biology Presentation by AbbyRaymundus Deaven GoanaNo ratings yet
- Sinclair Comp 2 Final Essay 5 1Document10 pagesSinclair Comp 2 Final Essay 5 1api-508910011No ratings yet
- Ad 8232Document8 pagesAd 8232umairNo ratings yet
- Adams-Stokes 1º Sintoma Febre ReumáticaDocument5 pagesAdams-Stokes 1º Sintoma Febre Reumáticacarvalho.tr4270No ratings yet
- The Circulatory System - Lecture STEMDocument50 pagesThe Circulatory System - Lecture STEMMargeory Calatan100% (1)
- I. Respiratory System: PhysiologyDocument2 pagesI. Respiratory System: PhysiologyVin BaniquedNo ratings yet
- Bio Grade 12 PhotosythesisDocument4 pagesBio Grade 12 PhotosythesisNardos DerejeNo ratings yet
- Ashiq Tutorials 2009Document24 pagesAshiq Tutorials 2009Sk ChaudhryNo ratings yet
- Apoptosis UsmleDocument2 pagesApoptosis Usmlelovelyc95100% (1)
- Physiology BCQsDocument25 pagesPhysiology BCQsReema KhanNo ratings yet
- NCP 1Document2 pagesNCP 1Momshie SyNo ratings yet
- Lower Respiratory DiseasesDocument137 pagesLower Respiratory DiseasesVictor StevenNo ratings yet