You might also like
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5813)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Game Theory and Animal Behavior (2000) by Lee Alan Dugatkin & Hudson Kern Reeve PDFDocument335 pagesGame Theory and Animal Behavior (2000) by Lee Alan Dugatkin & Hudson Kern Reeve PDFAnonymous gK48v9100% (1)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Genetic CentreDocument37 pagesGenetic CentreProf. Madhavan100% (3)
- STS Module 3 Section 3Document12 pagesSTS Module 3 Section 3Anthony HoNo ratings yet
- The Biogenesis and Función of PiRNAs in Human DiseasesDocument13 pagesThe Biogenesis and Función of PiRNAs in Human DiseasesArmando GBNo ratings yet
- Sistema Inmunitario y ESTRËSDocument15 pagesSistema Inmunitario y ESTRËSArmando GBNo ratings yet
- Hospital - Miocinas RevisiónDocument12 pagesHospital - Miocinas RevisiónArmando GBNo ratings yet
- Georganopoulou2021 Article AJourneyWithLGMDFromProteinAbnDocument23 pagesGeorganopoulou2021 Article AJourneyWithLGMDFromProteinAbnArmando GBNo ratings yet
- Multivariate Data Analysis For Omics: September 2-3 2008 Susanne Wiklund IID 1062Document45 pagesMultivariate Data Analysis For Omics: September 2-3 2008 Susanne Wiklund IID 1062homeNo ratings yet
- Motor Cortex Pyramidal ExtrapyramidalDocument22 pagesMotor Cortex Pyramidal ExtrapyramidalJOSE ARTURO LOZANO CRUZNo ratings yet
- Cambridge International AS & A Level: Biology 9700/33 October/November 2022Document6 pagesCambridge International AS & A Level: Biology 9700/33 October/November 2022mariakhan.educationNo ratings yet
- Datesheet of Unit Test - 3 (Iii-Xii) 2022-2023Document1 pageDatesheet of Unit Test - 3 (Iii-Xii) 2022-2023Techno HackerzNo ratings yet
- Worksheets Grammar 24Document1 pageWorksheets Grammar 24Emily castroNo ratings yet
- WDR-Weaver Dice RulebookDocument29 pagesWDR-Weaver Dice RulebookAzazelNo ratings yet
- Primer DesignDocument37 pagesPrimer DesignRyu- MikaNo ratings yet
- Gnbio1 Module 2Document35 pagesGnbio1 Module 2Ggggnek OhculNo ratings yet
- Pricelist 2023: Medical Laboratory-Hospital Equipment - Chemical SupplierDocument4 pagesPricelist 2023: Medical Laboratory-Hospital Equipment - Chemical SupplierAfriana SartinNo ratings yet
- Columbus Public School, Rudrapur: Syllabus For Session 2021-2022 Class - XIDocument5 pagesColumbus Public School, Rudrapur: Syllabus For Session 2021-2022 Class - XISsj rex 4No ratings yet
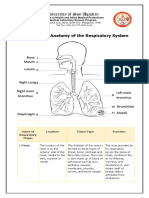
- Anatomy of The Respiratory SystemDocument4 pagesAnatomy of The Respiratory SystemMayden Grace GayatgayNo ratings yet
- Geriatric Nursing First ClassDocument45 pagesGeriatric Nursing First ClassPoonam RanaNo ratings yet
- Class 11 Biology Chapter 3 - Revision NotesDocument34 pagesClass 11 Biology Chapter 3 - Revision NotesAnubhav MamgainNo ratings yet
- Felipe - Assignment SPEC 211 PDFDocument1 pageFelipe - Assignment SPEC 211 PDFFrancene FelipeNo ratings yet
- Work For Sept. 23/24 Starts Here:: Unit 2: ProbabilityDocument52 pagesWork For Sept. 23/24 Starts Here:: Unit 2: ProbabilityRiddhiman PalNo ratings yet
- Item5 - Allsheng Products - pg19Document32 pagesItem5 - Allsheng Products - pg19RendraNo ratings yet
- RPS Biochemistry 2021Document13 pagesRPS Biochemistry 2021Naomi JuntakNo ratings yet
- YMMNO ProposalDocument2 pagesYMMNO ProposalYamon Min Nyunt OoNo ratings yet
- Grade 6: Subject Unit Title, Topics and Skills Unit Name, Topics and Skills Unit Name, Topics and SkillsDocument3 pagesGrade 6: Subject Unit Title, Topics and Skills Unit Name, Topics and Skills Unit Name, Topics and SkillsBagues ALit ManuabaNo ratings yet
- VRTS 111 Journal 1Document5 pagesVRTS 111 Journal 1Queen Angelique EsguerraNo ratings yet
- A New Species of The Genus Rhabdophis Fitzinger, 1843 (Squamata: Colubridae) From Guangdong Province, Southern ChinaDocument13 pagesA New Species of The Genus Rhabdophis Fitzinger, 1843 (Squamata: Colubridae) From Guangdong Province, Southern ChinaLĩnh NguyễnNo ratings yet
- Fish Capture and Volleyball NotesDocument5 pagesFish Capture and Volleyball NotesCalei MelioNo ratings yet
- CAMPANULA POLYMORPHA PublishedDocument4 pagesCAMPANULA POLYMORPHA PublishedAnca ManoleNo ratings yet
- Effects of Azospirillum IsolatesDocument8 pagesEffects of Azospirillum IsolatesOliver TalipNo ratings yet
- Mycology Systematics - Fungi ClassificationDocument14 pagesMycology Systematics - Fungi Classificationdlbook100% (1)
- Bacon 2006Document10 pagesBacon 2006Bryan Lloyd LauritoNo ratings yet
- Evolutionary Computation - A Unified ApproachDocument7 pagesEvolutionary Computation - A Unified ApproachSunena K.SNo ratings yet