You might also like
- Advanced Gynecologic Endoscopy: Edited by Atef DarwishDocument344 pagesAdvanced Gynecologic Endoscopy: Edited by Atef DarwishSandu Alexandra100% (1)
- Explicit Instruction: A Teaching Strategy in Reading, Writing, and Mathematics For Students With Learning DisabilitiesDocument3 pagesExplicit Instruction: A Teaching Strategy in Reading, Writing, and Mathematics For Students With Learning DisabilitiesKatherineNo ratings yet
- 4-7 The Law of Sines and The Law of Cosines PDFDocument40 pages4-7 The Law of Sines and The Law of Cosines PDFApple Vidal100% (1)
- Business Continuity Generaly Accepted Practices GAP v2.1 (Disaster Recovery Journal 2015) PDFDocument140 pagesBusiness Continuity Generaly Accepted Practices GAP v2.1 (Disaster Recovery Journal 2015) PDFducuhNo ratings yet
- Readings On The History and System of The Common Law - Roscoe PoundDocument646 pagesReadings On The History and System of The Common Law - Roscoe PoundpajorocNo ratings yet
- Inventory Valiuation Raw QueryDocument4 pagesInventory Valiuation Raw Querysatyanarayana NVSNo ratings yet
- Risus License Information PDFDocument1 pageRisus License Information PDFSam CorbenNo ratings yet
- ASTM D5895 - 2020 Tiempo SecadoDocument4 pagesASTM D5895 - 2020 Tiempo SecadoPablo OrtegaNo ratings yet
- Training Report PRASADDocument32 pagesTraining Report PRASADshekharazad_suman85% (13)
- A319/A320/A321 Technical Training Manual Mechanics / Electrics & Avionics Course 33 LightsDocument224 pagesA319/A320/A321 Technical Training Manual Mechanics / Electrics & Avionics Course 33 LightsAhmedHamdyElsaidy100% (3)
- Automation & Artificial Intelligence: Robots and Their ApplicationsDocument93 pagesAutomation & Artificial Intelligence: Robots and Their ApplicationsManal AndhereNo ratings yet
- Samples For Loanshyd-1Document3 pagesSamples For Loanshyd-1dpkraja100% (1)
- Ekinci BDocument4 pagesEkinci BreclinpharmaNo ratings yet
- Human Monocyte Response To Andean-Native Starch NanoparticlesDocument8 pagesHuman Monocyte Response To Andean-Native Starch NanoparticlesSol AngelNo ratings yet
- Sortase A Confers Protection Against Streptococcus Pneumoniae in MiceDocument5 pagesSortase A Confers Protection Against Streptococcus Pneumoniae in MiceRin ChanNo ratings yet
- Antimicrobial Agents and Chemotherapy-1998-Stuertz-277.fullDocument5 pagesAntimicrobial Agents and Chemotherapy-1998-Stuertz-277.fullCamila LaraNo ratings yet
- Morphological Changes of Klebsiella Pneumoniae in Response To Cefotaxime: A Scanning Electron Microscope StudyDocument4 pagesMorphological Changes of Klebsiella Pneumoniae in Response To Cefotaxime: A Scanning Electron Microscope StudyvanishivshettyNo ratings yet
- Zhu 2007Document9 pagesZhu 2007AnAn BanhGaoNo ratings yet
- High Prevalence of Methicillin-Resistant: Staphylococcus Haemolyticus Isolated From Commensals in Healthy AdultsDocument8 pagesHigh Prevalence of Methicillin-Resistant: Staphylococcus Haemolyticus Isolated From Commensals in Healthy AdultsElok NurfaiqohNo ratings yet
- II 1101006633Document10 pagesII 1101006633Rodrigo AlvesNo ratings yet
- Jones 2000Document12 pagesJones 2000choklletmrNo ratings yet
- 1 PBDocument5 pages1 PBRin ChanNo ratings yet
- Detection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.Document6 pagesDetection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.International Medical PublisherNo ratings yet
- 1 Alvarado2009Document10 pages1 Alvarado2009Eliza Aguilar MejíaNo ratings yet
- Antimicrob. Agents Chemother. 2009 Dai 393 400Document9 pagesAntimicrob. Agents Chemother. 2009 Dai 393 400Risma S PNo ratings yet
- 2003 - Journal of Endodontics - 29 - 4 - 240 - 243 - Induction of Cyclooxygenase 2 mRNA and Protein Expressio PDFDocument4 pages2003 - Journal of Endodontics - 29 - 4 - 240 - 243 - Induction of Cyclooxygenase 2 mRNA and Protein Expressio PDFNamrata SachdevaNo ratings yet
- Chhatwal 2002Document4 pagesChhatwal 2002Rin ChanNo ratings yet
- Virulence Factors in BacteriaDocument9 pagesVirulence Factors in BacteriaKESHAWN TYRONE SUTHERLANDNo ratings yet
- Functional and Phenotypic Changes in Monocytes From Patients With Tuberculosis Are Reversed With TreatmentDocument9 pagesFunctional and Phenotypic Changes in Monocytes From Patients With Tuberculosis Are Reversed With TreatmentCecile E EcheverríaNo ratings yet
- 2011 The LuxS-Dependent Quorum Sensing System Regulates Early Biofilm Formation by Streptococcus Pneumoniae Strain D39Document42 pages2011 The LuxS-Dependent Quorum Sensing System Regulates Early Biofilm Formation by Streptococcus Pneumoniae Strain D39hazyaNo ratings yet
- Cefotiam in VivoDocument10 pagesCefotiam in VivoVirus HepatitisCNo ratings yet
- BJSTR.MS.ID.005072Document6 pagesBJSTR.MS.ID.005072z97jxgjpwnNo ratings yet
- Mechanisms of Bacterial PathogenicityDocument10 pagesMechanisms of Bacterial PathogenicitymeliananitaNo ratings yet
- Antimicrob. Agents Chemother. 1995 Von Eiff 268 70Document3 pagesAntimicrob. Agents Chemother. 1995 Von Eiff 268 70Charisse Jan PercalesNo ratings yet
- Omaña-Molina Et Al., Type 2 DM BALB-c Mice Are More Susceptible To GAEDocument10 pagesOmaña-Molina Et Al., Type 2 DM BALB-c Mice Are More Susceptible To GAEIsmael CastelanNo ratings yet
- Influenza Type A Virus Neuraminidase Does Not Play A Role in Viral Entry, Replication, Assembly, or BuddingDocument8 pagesInfluenza Type A Virus Neuraminidase Does Not Play A Role in Viral Entry, Replication, Assembly, or BuddingNixie PeraltaNo ratings yet
- 1-s2.0-0168170286900080-mainDocument13 pages1-s2.0-0168170286900080-mainfalmazanNo ratings yet
- DocumentDocument10 pagesDocumentJayNo ratings yet
- Infect. Immun.-2001-Loukas-3646-51Document6 pagesInfect. Immun.-2001-Loukas-3646-51Herick MullerNo ratings yet
- Analysis of Inhibitory Propensity and Percentage Inhibition of Tecoma Stans and PlectranthusDocument6 pagesAnalysis of Inhibitory Propensity and Percentage Inhibition of Tecoma Stans and PlectranthusSabrina JonesNo ratings yet
- Suay 2000 ATMB BasidyoDocument11 pagesSuay 2000 ATMB Basidyoinositol66No ratings yet
- Mechanism of Bacterial PathogenicityDocument10 pagesMechanism of Bacterial PathogenicityBetty Debora KlaraNo ratings yet
- 511 jmm000460Document6 pages511 jmm000460Nur LaylaNo ratings yet
- Metronidazole Induces Programmed Cell Death in Blastocystis hominisDocument11 pagesMetronidazole Induces Programmed Cell Death in Blastocystis hominisLiezle GargolesNo ratings yet
- Emetine Produce Entamoeba Histolytica Death by IndDocument6 pagesEmetine Produce Entamoeba Histolytica Death by IndJota CarlosNo ratings yet
- Acidosalicilico ExossomaDocument7 pagesAcidosalicilico ExossomaRenato GranadoNo ratings yet
- 338111v4 FullDocument37 pages338111v4 FullIlincaNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsDiego TulcanNo ratings yet
- Replacement For Bacteriophage Typing of Pulsed-Field Gel Electrophoresis As ADocument6 pagesReplacement For Bacteriophage Typing of Pulsed-Field Gel Electrophoresis As ANandakurniasariNo ratings yet
- Ijmr 5 2 179 183Document5 pagesIjmr 5 2 179 183Abhay KumarNo ratings yet
- Veterinary Immunology and ImmunopathologyDocument6 pagesVeterinary Immunology and ImmunopathologySheikh FirdousNo ratings yet
- Salmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008Document8 pagesSalmonella Typhi: From A Human Pathogen To A Vaccine Vector: Cellular & Molecular Immunology May 2008noordin MukasaNo ratings yet
- tmp7DDD TMPDocument6 pagestmp7DDD TMPFrontiersNo ratings yet
- Pathogenesis in Tuberculosis: Transcriptomic Approaches To Unraveling Virulence Mechanisms and Finding New Drug TargetsDocument23 pagesPathogenesis in Tuberculosis: Transcriptomic Approaches To Unraveling Virulence Mechanisms and Finding New Drug Targetskinta translateNo ratings yet
- Artigo RubensDocument8 pagesArtigo RubensbuissaNo ratings yet
- CSOM Etiological Agents and Antibiotic SensitivityDocument4 pagesCSOM Etiological Agents and Antibiotic Sensitivitywilson wijayaNo ratings yet
- Mscano, Layout Editor, Jidc-011-0513Document4 pagesMscano, Layout Editor, Jidc-011-0513dary.podoliak03No ratings yet
- Infections of M. PneumoniaeDocument18 pagesInfections of M. PneumoniaeAndrés Nicolás Beltrán SeguraNo ratings yet
- Isolation and Characterization of Methicillin-Resistant Strains From Nares of Nurses and Their GownsDocument5 pagesIsolation and Characterization of Methicillin-Resistant Strains From Nares of Nurses and Their Gownskhaled msalbiNo ratings yet
- Modified P.aerugenosaDocument21 pagesModified P.aerugenosaranjitsinghNo ratings yet
- Modified P.aerugenosaDocument21 pagesModified P.aerugenosaranjitsinghNo ratings yet
- Effects of Sodium Ascorbate (Vitamin C) and 2-Methyl-1,4-Naphthoquinone (Vitamin K3) Treatment On Human Tumor Cell Growth in VitroDocument6 pagesEffects of Sodium Ascorbate (Vitamin C) and 2-Methyl-1,4-Naphthoquinone (Vitamin K3) Treatment On Human Tumor Cell Growth in VitroUtopiaawaits100% (1)
- 3127-Article Text-25112-1-10-20131114Document8 pages3127-Article Text-25112-1-10-20131114guizah nourelhoudaNo ratings yet
- Kvir 15 2301246Document17 pagesKvir 15 2301246Rodrigo AlvesNo ratings yet
- AnthraxDocument12 pagesAnthraxAvi Verma100% (1)
- Microbial Pathogenesis: SciencedirectDocument6 pagesMicrobial Pathogenesis: SciencedirectioanaNo ratings yet
- Effect of Taxol From Pestalotiopsis Mangiferae On A549 Cells-In Vitro StudyDocument9 pagesEffect of Taxol From Pestalotiopsis Mangiferae On A549 Cells-In Vitro StudyLizbethNo ratings yet
- tmpBF53 TMPDocument6 pagestmpBF53 TMPFrontiersNo ratings yet
- Modeled Microgravity Conditions Suppress Innate Macrophage and Lymphocytic Responses To Common Mitogens and MycobacteriumDocument9 pagesModeled Microgravity Conditions Suppress Innate Macrophage and Lymphocytic Responses To Common Mitogens and MycobacteriumAtrocitus RedNo ratings yet
- The Veterinary Journal: Jun Zhang, Gui-Hong Zhang, Lin Yang, Ren Huang, Yu Zhang, Kun Jia, Wen Yuan, Shou-Jun LiDocument4 pagesThe Veterinary Journal: Jun Zhang, Gui-Hong Zhang, Lin Yang, Ren Huang, Yu Zhang, Kun Jia, Wen Yuan, Shou-Jun LiEdgar DíazNo ratings yet
- Detection of Major Facilitator Superfamily (MFS) Transporter in Enterobacteriaceae Isolated From Chicken (Rizal Et Al., 2022)Document6 pagesDetection of Major Facilitator Superfamily (MFS) Transporter in Enterobacteriaceae Isolated From Chicken (Rizal Et Al., 2022)Syaiful RizalNo ratings yet
- Small Molecular Immunomodifiers of Microbial Origin: Fundamental and Clinical Studies of BestatinFrom EverandSmall Molecular Immunomodifiers of Microbial Origin: Fundamental and Clinical Studies of BestatinHamao UmezawaNo ratings yet
- Lipid Profiles with Diagnoses Under 40 CharactersDocument21 pagesLipid Profiles with Diagnoses Under 40 CharactersSandu AlexandraNo ratings yet
- Joi 150056Document9 pagesJoi 150056Rendy SusantoNo ratings yet
- 1 s2.0 S1214021X14600260 MainDocument10 pages1 s2.0 S1214021X14600260 MainSandu AlexandraNo ratings yet
- Poillucci2017 Article LaparoscopicAppendectomyVsAntiDocument10 pagesPoillucci2017 Article LaparoscopicAppendectomyVsAntiSandu AlexandraNo ratings yet
- Allievi2017 Article AcuteAppendicitisStillASurgicaDocument9 pagesAllievi2017 Article AcuteAppendicitisStillASurgicaSandu AlexandraNo ratings yet
- 1 s2.0 S1479666X17300562 MainDocument12 pages1 s2.0 S1479666X17300562 MainSandu AlexandraNo ratings yet
- Thyroid Surgery Throughout History: EditorialDocument4 pagesThyroid Surgery Throughout History: EditorialSandu AlexandraNo ratings yet
- 1 s2.0 S0196655316001693 MainDocument3 pages1 s2.0 S0196655316001693 MainSandu AlexandraNo ratings yet
- The Current Protocols in The Management of Hepatic Hydatid DiseaseDocument8 pagesThe Current Protocols in The Management of Hepatic Hydatid DiseaseSandu AlexandraNo ratings yet
- Butt 2019Document5 pagesButt 2019Sandu AlexandraNo ratings yet
- Percutaneous Treatment of Liver Hydatid Cysts in 190 Patients: A Retrospective StudyDocument9 pagesPercutaneous Treatment of Liver Hydatid Cysts in 190 Patients: A Retrospective StudySandu AlexandraNo ratings yet
- Surgical Techniques For Management Hepatic Hydatid Cysts in Fallujah Teaching Hospital: A Prospective StudyDocument4 pagesSurgical Techniques For Management Hepatic Hydatid Cysts in Fallujah Teaching Hospital: A Prospective StudySandu AlexandraNo ratings yet
- E00261-16 FullDocument12 pagesE00261-16 FullSandu AlexandraNo ratings yet
- 1 s2.0 S1198743X14613203 MainDocument11 pages1 s2.0 S1198743X14613203 MainSandu AlexandraNo ratings yet
- Operative Treatment of Hepatic Hydatid Cysts: A Single Center ExperienceDocument6 pagesOperative Treatment of Hepatic Hydatid Cysts: A Single Center ExperienceSandu AlexandraNo ratings yet
- Pneumococcus: A Deadly but Declining Infectious DiseaseDocument8 pagesPneumococcus: A Deadly but Declining Infectious DiseaseSandu AlexandraNo ratings yet
- Major Liver Resection For Recurrent Hydatid Cyst of The Liver After Suboptimal TreatmentDocument6 pagesMajor Liver Resection For Recurrent Hydatid Cyst of The Liver After Suboptimal TreatmentSandu AlexandraNo ratings yet
- Comparison of Long-Term Results of Percutaneous Treatment Techniques For Hepatic Cystic Echinococcosis Types 2 and 3bDocument7 pagesComparison of Long-Term Results of Percutaneous Treatment Techniques For Hepatic Cystic Echinococcosis Types 2 and 3bSandu AlexandraNo ratings yet
- Successful Percutaneous Treatment of Extrahepatic Cystic Echinococcosis Through PAIR and Single Puncture Catheter TechniquesDocument7 pagesSuccessful Percutaneous Treatment of Extrahepatic Cystic Echinococcosis Through PAIR and Single Puncture Catheter TechniquesSandu AlexandraNo ratings yet
- Treatment of Intra-Abdominal Hydatid Cyst Using Modified PAIR Technique: Single Center ExperiencesDocument5 pagesTreatment of Intra-Abdominal Hydatid Cyst Using Modified PAIR Technique: Single Center ExperiencesSandu AlexandraNo ratings yet
- Arab Journal of GastroenterologyDocument9 pagesArab Journal of GastroenterologySandu AlexandraNo ratings yet
- Factors Affecting The Choice of Treatment in Hepatic Hydatid Cyst SurgeryDocument7 pagesFactors Affecting The Choice of Treatment in Hepatic Hydatid Cyst SurgerySandu AlexandraNo ratings yet
- Clinicalinvestigation Non-VascularinterventionsDocument7 pagesClinicalinvestigation Non-VascularinterventionsSandu AlexandraNo ratings yet
- A Novel Modified PAIR Technique Using A Trocar Catheter For Percutaneous Treatment of Liver Hydatid Cysts: A Six-Year ExperienceDocument5 pagesA Novel Modified PAIR Technique Using A Trocar Catheter For Percutaneous Treatment of Liver Hydatid Cysts: A Six-Year ExperienceSandu AlexandraNo ratings yet
- Hepatic Hydatid Cyst Misdiagnosed As Simple Cyst: A Case ReportDocument6 pagesHepatic Hydatid Cyst Misdiagnosed As Simple Cyst: A Case ReportSandu AlexandraNo ratings yet
- 8Document8 pages8Sandu AlexandraNo ratings yet
- Ovarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentDocument7 pagesOvarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentSandu AlexandraNo ratings yet
- Case ReportDocument5 pagesCase ReportNurRohmanNo ratings yet
- Fit Friend Business Game StrategiesDocument7 pagesFit Friend Business Game StrategiesSanchit AggarwalNo ratings yet
- Power Fresh 16Document3 pagesPower Fresh 16M. Ardi PrediyanaNo ratings yet
- Laser Beam Machining (LBM)Document10 pagesLaser Beam Machining (LBM)RAMAKANT RANANo ratings yet
- Gec-Art Art Appreciation: Course Code: Course Title: Course DescriptionsDocument14 pagesGec-Art Art Appreciation: Course Code: Course Title: Course Descriptionspoleene de leonNo ratings yet
- Performance Evaluation of Root Crop HarvestersDocument15 pagesPerformance Evaluation of Root Crop HarvestersIJERDNo ratings yet
- Workbook. Unit 3. Exercises 5 To 9. RESPUESTASDocument3 pagesWorkbook. Unit 3. Exercises 5 To 9. RESPUESTASRosani GeraldoNo ratings yet
- MT Series User Manual MT4YDocument28 pagesMT Series User Manual MT4YDhani Aristyawan SimangunsongNo ratings yet
- Yatendra Kumar Sharma ResumeDocument3 pagesYatendra Kumar Sharma ResumeDheeraj SharmaNo ratings yet
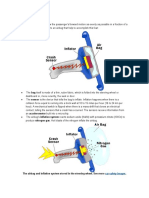
- Airbag Inflation: The Airbag and Inflation System Stored in The Steering Wheel. See MoreDocument5 pagesAirbag Inflation: The Airbag and Inflation System Stored in The Steering Wheel. See MoreShivankur HingeNo ratings yet
- Ebooks vs Traditional Books: An AnalysisDocument10 pagesEbooks vs Traditional Books: An AnalysisLOVENo ratings yet
- Basic of Seismic RefractionDocument43 pagesBasic of Seismic Refractionfitriah wulandariNo ratings yet
- D-2 UTEP Currey Final ReportDocument23 pagesD-2 UTEP Currey Final ReporthmcNo ratings yet
- Eliminator ® Liquid Line Fi Lter Driers: DML and DCLDocument12 pagesEliminator ® Liquid Line Fi Lter Driers: DML and DCLbracaNSNo ratings yet
- India: Soil Types, Problems & Conservation: Dr. SupriyaDocument25 pagesIndia: Soil Types, Problems & Conservation: Dr. SupriyaManas KaiNo ratings yet
- LEEA-030.2c2 Certificate of Thorough Examination (Multiple Items) (Overseas) (Dev)Document1 pageLEEA-030.2c2 Certificate of Thorough Examination (Multiple Items) (Overseas) (Dev)GaniyuNo ratings yet
- Reference Interval For CeruloplasminDocument3 pagesReference Interval For CeruloplasminGawri AbeyNo ratings yet
- Ic T7HDocument36 pagesIc T7HCarlos GaiarinNo ratings yet
- Stockholm Acc A300 600 2278Document164 pagesStockholm Acc A300 600 2278tugayyoungNo ratings yet
- Aptamers in HIV Research Diagnosis and TherapyDocument11 pagesAptamers in HIV Research Diagnosis and TherapymikiNo ratings yet