You might also like
- Attractor Networks: Fundamentals and Applications in Computational NeuroscienceFrom EverandAttractor Networks: Fundamentals and Applications in Computational NeuroscienceNo ratings yet
- Zhang-Zhao2019 Article FluorescenceMicroscopyImageClaDocument12 pagesZhang-Zhao2019 Article FluorescenceMicroscopyImageClaMahaNo ratings yet
- IET Image Processing - 2023 - Zhou - Accurately 3D Neuron Localization Using 2D Conv LSTM Super Resolution SegmentationDocument14 pagesIET Image Processing - 2023 - Zhou - Accurately 3D Neuron Localization Using 2D Conv LSTM Super Resolution SegmentationAnu s sNo ratings yet
- ISBI2019 HuiDocument5 pagesISBI2019 Huizj xiaoNo ratings yet
- A Literature Review On Pseudotime Estimation Using Single-Cell DataDocument4 pagesA Literature Review On Pseudotime Estimation Using Single-Cell DataInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Deep Learning With A Classifier SystemDocument9 pagesDeep Learning With A Classifier SystemlarisszaNo ratings yet
- Using Deep Correlation Features To Define The Meta Style of Cell Images For ClassificationDocument4 pagesUsing Deep Correlation Features To Define The Meta Style of Cell Images For ClassificationneigeNo ratings yet
- 1 s2.0 S2667305323000121 MainDocument10 pages1 s2.0 S2667305323000121 MainSumeet MitraNo ratings yet
- Hover-Net: Simultaneous Segmentation and Classification of Nuclei in Multi-Tissue Histology ImagesDocument19 pagesHover-Net: Simultaneous Segmentation and Classification of Nuclei in Multi-Tissue Histology ImagesAhmed WaelNo ratings yet
- Anomaly Detection On Attributed NetworksDocument15 pagesAnomaly Detection On Attributed NetworksbarracudaNo ratings yet
- Axondeepseg: Automatic Axon and Myelin Segmentation From Microscopy Data Using Convolutional Neural NetworksDocument11 pagesAxondeepseg: Automatic Axon and Myelin Segmentation From Microscopy Data Using Convolutional Neural Networksmarko CavdarNo ratings yet
- Aip/123 QedDocument49 pagesAip/123 QedAlessio GagliardiNo ratings yet
- Corresponding AuthorDocument6 pagesCorresponding AuthorArs Arifur RahmanNo ratings yet
- Capsule NetworkDocument8 pagesCapsule NetworkMalia HaleNo ratings yet
- Modeling Systems With Internal State Using EvolinoDocument9 pagesModeling Systems With Internal State Using EvolinoRENI YUNITA 227056004No ratings yet
- Biomedical Research PaperDocument7 pagesBiomedical Research PaperSathvik ShenoyNo ratings yet
- Quantum Capsule NetworksDocument18 pagesQuantum Capsule NetworksRidho AkademikNo ratings yet
- A Deep Learning-Based Algorithm For 2-D Cell Segmentation in Microscopy ImagesDocument11 pagesA Deep Learning-Based Algorithm For 2-D Cell Segmentation in Microscopy ImagesIgnasi AlemanyNo ratings yet
- 251 SubmissionDocument6 pages251 Submissionsharwari solapurNo ratings yet
- Zhong Et Al. - 2017 - Learning To Diversify Deep Belief Networks For Hyperspectral Image ClassificationDocument15 pagesZhong Et Al. - 2017 - Learning To Diversify Deep Belief Networks For Hyperspectral Image ClassificationEwerton DuarteNo ratings yet
- Visualizing Hierarchies in scRNA-seq Data Using A Density Tree-Biased AutoencoderDocument18 pagesVisualizing Hierarchies in scRNA-seq Data Using A Density Tree-Biased AutoencoderRomina TurcoNo ratings yet
- An Improved Target Detection Algorithm Based On EfficientnetDocument10 pagesAn Improved Target Detection Algorithm Based On EfficientnetNeha BhatiNo ratings yet
- A Novel Cell Segmentation Method and Cell Phase IdDocument17 pagesA Novel Cell Segmentation Method and Cell Phase IdSleem KsNo ratings yet
- The Emergence and Promise of Single-Cell Temporal-Omics ApproachesDocument9 pagesThe Emergence and Promise of Single-Cell Temporal-Omics ApproachesFrancisco Javier BaculimaNo ratings yet
- Unsupervised Learning Using Back Propagation in Neural NetworksDocument4 pagesUnsupervised Learning Using Back Propagation in Neural NetworksPrince GurajenaNo ratings yet
- Image Classification With Feed-Forward Neural Networks: Meller, Matula and ChłądDocument7 pagesImage Classification With Feed-Forward Neural Networks: Meller, Matula and ChłądSoumini PortfolioNo ratings yet
- From Deep Learning To Mechanistic Understanding in Neuroscience: The Structure of Retinal PredictionDocument15 pagesFrom Deep Learning To Mechanistic Understanding in Neuroscience: The Structure of Retinal Predictiontiva salvationNo ratings yet
- Journal Pcbi 1009879Document33 pagesJournal Pcbi 1009879RmzNo ratings yet
- Improved Automatic Detection and Segmentation of Cell Nuclei in Histopathology ImagesDocument12 pagesImproved Automatic Detection and Segmentation of Cell Nuclei in Histopathology ImagesEgor FenkoNo ratings yet
- A New Convolutional Neural Network Architecture For Automatic Detection of Brain Tumors in Magnetic Resonance Imaging ImagesDocument8 pagesA New Convolutional Neural Network Architecture For Automatic Detection of Brain Tumors in Magnetic Resonance Imaging Imagessujay KumarrunNo ratings yet
- Bio Technologia 2013943Document17 pagesBio Technologia 2013943Nam Hoàng VănNo ratings yet
- A D O - C N N: Nomaly Etection Using NE Lass Eural EtworksDocument13 pagesA D O - C N N: Nomaly Etection Using NE Lass Eural Etworksdipak786No ratings yet
- See Elegans: Simple-To-Use, Accurate, and Automatic 3D Detection of Neural Activity From Densely Packed NeuronsDocument21 pagesSee Elegans: Simple-To-Use, Accurate, and Automatic 3D Detection of Neural Activity From Densely Packed NeuronsNikhil ChoudharyNo ratings yet
- Learning A Complete Image Indexing PipelineDocument10 pagesLearning A Complete Image Indexing Pipelinesubi5No ratings yet
- Symbolic Models For DLDocument25 pagesSymbolic Models For DLzentropiaNo ratings yet
- Print 5Document17 pagesPrint 5Nepsis ElvețiaNo ratings yet
- Classification of MR Brain Images For Detection of Tumor With Transfer Learning From Pre-Trained CNN ModelsDocument4 pagesClassification of MR Brain Images For Detection of Tumor With Transfer Learning From Pre-Trained CNN Modelseshwari2000No ratings yet
- 1 s2.0 S0169260723005904 MainDocument10 pages1 s2.0 S0169260723005904 MainMohamed TahirNo ratings yet
- Mca 27 00022 v2Document35 pagesMca 27 00022 v2mx_maziNo ratings yet
- ATSSDocument10 pagesATSSMateus RibeiroNo ratings yet
- High-Speed Fluorescence Image - Enabled Cell Sorting: Research TechnologyDocument7 pagesHigh-Speed Fluorescence Image - Enabled Cell Sorting: Research Technologyjessica hernandezNo ratings yet
- IJSRDV8I110168Document5 pagesIJSRDV8I110168Chaudhary MuskaanNo ratings yet
- Paper 2Document13 pagesPaper 2Asmi SinhaNo ratings yet
- High Speed Tracking With Kernelized Correlation FiltersDocument14 pagesHigh Speed Tracking With Kernelized Correlation FiltersLarry CocaNo ratings yet
- Local Aggregation For Unsupervised Learning of Visual EmbeddingsDocument13 pagesLocal Aggregation For Unsupervised Learning of Visual EmbeddingsKamalesh SNo ratings yet
- HardAttentionNetforRetinaVesselSegmentation FinalDocument13 pagesHardAttentionNetforRetinaVesselSegmentation Finalyt HehkkeNo ratings yet
- Tmi 2015 2457891Document11 pagesTmi 2015 2457891Osama Ayad KadimNo ratings yet
- Applsci 13 12977Document21 pagesApplsci 13 12977gopineedivigneshNo ratings yet
- Chen 2018Document14 pagesChen 2018muraliNo ratings yet
- 1807 10584fhwfDocument6 pages1807 10584fhwfTiến Việt PhạmNo ratings yet
- Zhang 2018Document12 pagesZhang 2018Reza NugrahaNo ratings yet
- RACE-Net: A Recurrent Neural Network For Biomedical Image SegmentationDocument12 pagesRACE-Net: A Recurrent Neural Network For Biomedical Image SegmentationSnehaNo ratings yet
- Developmental Stereo Emergence of Disparity Preference in Models of The Visual Cortex-ujBDocument15 pagesDevelopmental Stereo Emergence of Disparity Preference in Models of The Visual Cortex-ujBmohmmaed khalidNo ratings yet
- GRUUNetDocument12 pagesGRUUNetAshish SinghNo ratings yet
- RetinaDocument23 pagesRetinaPeruru Famida NajumunNo ratings yet
- A Detailed Look at CNN-based Approaches in Facial Landmark DetectionDocument9 pagesA Detailed Look at CNN-based Approaches in Facial Landmark DetectionRanaBilalShahidNo ratings yet
- Paper 4Document11 pagesPaper 4Mapache CientíficoNo ratings yet
- Pattern Recognition: Muhammad Iqbal, Syed S. Naqvi, Will N. Browne, Christopher Hollitt, Mengjie ZhangDocument15 pagesPattern Recognition: Muhammad Iqbal, Syed S. Naqvi, Will N. Browne, Christopher Hollitt, Mengjie ZhangTamara SalehNo ratings yet
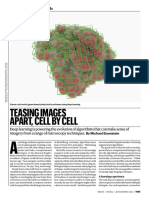
- Teasing Images Apart, Cell by CellDocument3 pagesTeasing Images Apart, Cell by Cellbiomarinium76No ratings yet
- Ridge Regression Neural Network For Pediatric Bone Age AssessmentDocument10 pagesRidge Regression Neural Network For Pediatric Bone Age AssessmentAliza MustafaNo ratings yet
- Microsoft Azure Infrastructure Services for Architects: Designing Cloud SolutionsFrom EverandMicrosoft Azure Infrastructure Services for Architects: Designing Cloud SolutionsNo ratings yet
- The Compete Ccna 200-301 Study Guide: Network Engineering EditionFrom EverandThe Compete Ccna 200-301 Study Guide: Network Engineering EditionRating: 5 out of 5 stars5/5 (4)
- Cybersecurity: A Simple Beginner’s Guide to Cybersecurity, Computer Networks and Protecting Oneself from Hacking in the Form of Phishing, Malware, Ransomware, and Social EngineeringFrom EverandCybersecurity: A Simple Beginner’s Guide to Cybersecurity, Computer Networks and Protecting Oneself from Hacking in the Form of Phishing, Malware, Ransomware, and Social EngineeringRating: 5 out of 5 stars5/5 (40)
- Cybersecurity: The Beginner's Guide: A comprehensive guide to getting started in cybersecurityFrom EverandCybersecurity: The Beginner's Guide: A comprehensive guide to getting started in cybersecurityRating: 5 out of 5 stars5/5 (2)
- Computer Networking: The Complete Beginner's Guide to Learning the Basics of Network Security, Computer Architecture, Wireless Technology and Communications Systems (Including Cisco, CCENT, and CCNA)From EverandComputer Networking: The Complete Beginner's Guide to Learning the Basics of Network Security, Computer Architecture, Wireless Technology and Communications Systems (Including Cisco, CCENT, and CCNA)Rating: 4 out of 5 stars4/5 (4)
- FTTx Networks: Technology Implementation and OperationFrom EverandFTTx Networks: Technology Implementation and OperationRating: 5 out of 5 stars5/5 (1)
- Evaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsFrom EverandEvaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsRating: 5 out of 5 stars5/5 (1)
- AWS Certified Solutions Architect Study Guide: Associate SAA-C01 ExamFrom EverandAWS Certified Solutions Architect Study Guide: Associate SAA-C01 ExamRating: 4 out of 5 stars4/5 (1)
- ITIL® 4 Create, Deliver and Support (CDS): Your companion to the ITIL 4 Managing Professional CDS certificationFrom EverandITIL® 4 Create, Deliver and Support (CDS): Your companion to the ITIL 4 Managing Professional CDS certificationRating: 5 out of 5 stars5/5 (2)
- AWS Certified Solutions Architect Study Guide: Associate SAA-C02 ExamFrom EverandAWS Certified Solutions Architect Study Guide: Associate SAA-C02 ExamNo ratings yet
- Open Radio Access Network (O-RAN) Systems Architecture and DesignFrom EverandOpen Radio Access Network (O-RAN) Systems Architecture and DesignNo ratings yet
- Network+ Study Guide & Practice ExamsFrom EverandNetwork+ Study Guide & Practice ExamsRating: 4.5 out of 5 stars4.5/5 (5)
- The Windows Command Line Beginner's Guide: Second EditionFrom EverandThe Windows Command Line Beginner's Guide: Second EditionRating: 4 out of 5 stars4/5 (4)
- Amazon Web Services (AWS) Interview Questions and AnswersFrom EverandAmazon Web Services (AWS) Interview Questions and AnswersRating: 4.5 out of 5 stars4.5/5 (3)
- CWNA Certified Wireless Network Administrator Study Guide: Exam CWNA-108From EverandCWNA Certified Wireless Network Administrator Study Guide: Exam CWNA-108No ratings yet
- Microsoft Certified Azure Fundamentals Study Guide: Exam AZ-900From EverandMicrosoft Certified Azure Fundamentals Study Guide: Exam AZ-900No ratings yet
- Hacking Network Protocols: Complete Guide about Hacking, Scripting and Security of Computer Systems and Networks.From EverandHacking Network Protocols: Complete Guide about Hacking, Scripting and Security of Computer Systems and Networks.Rating: 5 out of 5 stars5/5 (2)
- ITIL 4 : Drive Stakeholder Value: Reference and study guideFrom EverandITIL 4 : Drive Stakeholder Value: Reference and study guideNo ratings yet
- ITIL® 4 Drive Stakeholder Value (DSV): Your companion to the ITIL 4 Managing Professional DSV certificationFrom EverandITIL® 4 Drive Stakeholder Value (DSV): Your companion to the ITIL 4 Managing Professional DSV certificationNo ratings yet
- Hacking: A Beginners Guide To Your First Computer Hack; Learn To Crack A Wireless Network, Basic Security Penetration Made Easy and Step By Step Kali LinuxFrom EverandHacking: A Beginners Guide To Your First Computer Hack; Learn To Crack A Wireless Network, Basic Security Penetration Made Easy and Step By Step Kali LinuxRating: 4.5 out of 5 stars4.5/5 (67)