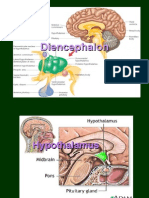
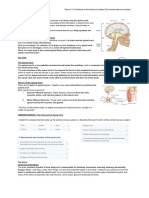
You might also like
- Pessoa2010 Emotion Processing and The AmydgalaDocument10 pagesPessoa2010 Emotion Processing and The AmydgalaMaria CordeiroNo ratings yet
- 174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperienceDocument20 pages174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperiencejuannnnNo ratings yet
- Brain Activity: Making HowDocument3 pagesBrain Activity: Making Howkickinghorse892No ratings yet
- Antrobus 1991Document26 pagesAntrobus 1991Vítor Gama (Vitium)No ratings yet
- Three Anti Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesThree Anti Correlated Neuronal Networks Managing Brain ActivitydgavrileNo ratings yet
- Cirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesCirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDanaNo ratings yet
- RepresentationDocument12 pagesRepresentationMike AngeluxNo ratings yet
- The Global Landscape of Cognition - Hierarchical Aggregation As An Organizational Principle of Human Cortical Networks and Functions Srep18112Document18 pagesThe Global Landscape of Cognition - Hierarchical Aggregation As An Organizational Principle of Human Cortical Networks and Functions Srep18112Peter PetroffNo ratings yet
- Wagner 1998Document5 pagesWagner 1998kickinghorse892No ratings yet
- Pearson 2019 AphantasiaDocument11 pagesPearson 2019 Aphantasiazhongkailin739No ratings yet
- Menon Short Course 10Document11 pagesMenon Short Course 10Kaan YılmazNo ratings yet
- 9 - miller2001CCDocument41 pages9 - miller2001CCFrancisco Ahumada MéndezNo ratings yet
- Empathy Circuits: Haakon G Engen and Tania SingerDocument8 pagesEmpathy Circuits: Haakon G Engen and Tania SingersupatraNo ratings yet
- Three Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewDocument9 pagesThree Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewVioleta-Nella MocanuNo ratings yet
- TMP 5 FADocument12 pagesTMP 5 FAFrontiersNo ratings yet
- Are Virtual Photons The Elementary Carriers of ConsciousnessDocument21 pagesAre Virtual Photons The Elementary Carriers of Consciousnessm:mNo ratings yet
- (TEO) CPF (Miller y Cogen, 2001)Document41 pages(TEO) CPF (Miller y Cogen, 2001)Eminencia Garcia PerezNo ratings yet
- The Neural Correlates of Conscious VisionDocument14 pagesThe Neural Correlates of Conscious VisionKarla ValderramaNo ratings yet
- Autonomous Cognitive Robots Need Emotional Modulations: Introducing The eMODUL ModelDocument10 pagesAutonomous Cognitive Robots Need Emotional Modulations: Introducing The eMODUL ModelJuan RomayNo ratings yet
- Neurobiology RelatedDocument9 pagesNeurobiology RelateddgavrileNo ratings yet
- Normal and Pathological Oscillatory Communication in The BrainDocument12 pagesNormal and Pathological Oscillatory Communication in The BrainDiegoNo ratings yet
- Biological MotionDocument14 pagesBiological MotionchysaNo ratings yet
- The Dorsal Frontoparietal Network: A Core System For Emulated ActionDocument11 pagesThe Dorsal Frontoparietal Network: A Core System For Emulated ActionDa CuNo ratings yet
- Using Goal-Driven Deep Learning Models To Understand The Sensory CortexDocument10 pagesUsing Goal-Driven Deep Learning Models To Understand The Sensory CortexRajsuryan SinghNo ratings yet
- Gray Et Al. (2002) .Integration of Emotion and Cognition in The Lateral Prefrontal CortexDocument6 pagesGray Et Al. (2002) .Integration of Emotion and Cognition in The Lateral Prefrontal CortexJosh CarrNo ratings yet
- Natural Behavior Is The Language of The BrainDocument12 pagesNatural Behavior Is The Language of The Brain테라테라No ratings yet
- Buzsaky y Moser 2013 Theta Entorrinal CXDocument22 pagesBuzsaky y Moser 2013 Theta Entorrinal CXCindy ZuritaNo ratings yet
- Amodio & Frith 2006 Meeting of Minds - The Medial Frontal Cortex and Social Cognition PDFDocument10 pagesAmodio & Frith 2006 Meeting of Minds - The Medial Frontal Cortex and Social Cognition PDFJen CuzzNo ratings yet
- Spunt (2011) JOCNDocument12 pagesSpunt (2011) JOCNBUBUSNo ratings yet
- Schizophrenia and The Mirror System: An Essay: Michael A. Arbib, T. Nathan MundhenkDocument13 pagesSchizophrenia and The Mirror System: An Essay: Michael A. Arbib, T. Nathan MundhenkMonica MargeNo ratings yet
- Rede Fining The Role of Limbic Areas in Cortical Processing: OpinionDocument11 pagesRede Fining The Role of Limbic Areas in Cortical Processing: OpinionMontserratNo ratings yet
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Document20 pagesGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNo ratings yet
- Poster DANUBE 2005 - Working Memory and EEG CoherenceDocument2 pagesPoster DANUBE 2005 - Working Memory and EEG Coherencesimon brežanNo ratings yet
- NIH Public Access: Author ManuscriptDocument14 pagesNIH Public Access: Author Manuscriptece142No ratings yet
- Understanding Emotions in Others: Mirror Neuron Dysfunction in Children With Autism Spectrum DisordersDocument3 pagesUnderstanding Emotions in Others: Mirror Neuron Dysfunction in Children With Autism Spectrum DisordersJohnNo ratings yet
- 1 s2.0 S0959438818300813 MainDocument7 pages1 s2.0 S0959438818300813 MainLilianne Mbengani LaranjeiraNo ratings yet
- Critchley Et Al 2004 NatNeurosci - Neural Systems Supporting Interoceptive AwarenessDocument7 pagesCritchley Et Al 2004 NatNeurosci - Neural Systems Supporting Interoceptive AwarenessAlanFerminNo ratings yet
- Nihms 474787Document7 pagesNihms 474787ClaraCalvoTorresNo ratings yet
- Frontal Lobe SyndromesDocument7 pagesFrontal Lobe SyndromespiedrahitasaNo ratings yet
- Fronto-Parietal Mirror Neuron System Modeling: Visuospatial Transformations Support Imitation Learning Independently of Imitator PerspectiveDocument21 pagesFronto-Parietal Mirror Neuron System Modeling: Visuospatial Transformations Support Imitation Learning Independently of Imitator PerspectiveNéstorNo ratings yet
- Department of Instrumentation EngineeringDocument28 pagesDepartment of Instrumentation EngineeringMahi KatreNo ratings yet
- Neuroimaging Studies of The Cerebellum LanguageDocument8 pagesNeuroimaging Studies of The Cerebellum Languagevikhen0712No ratings yet
- The Mirror Neuron System: Luigi Cattaneo, MD Giacomo Rizzolatti, MDDocument4 pagesThe Mirror Neuron System: Luigi Cattaneo, MD Giacomo Rizzolatti, MDece142No ratings yet
- Affective Mapping An Activation Likelihood Estimation (ALE) Meta-AnalysisDocument12 pagesAffective Mapping An Activation Likelihood Estimation (ALE) Meta-Analysisselina hahnNo ratings yet
- The Sensory Neurons of Touch (Abraira & Ginty, 2013)Document41 pagesThe Sensory Neurons of Touch (Abraira & Ginty, 2013)GokushimakNo ratings yet
- 1988 Perspectives On Cognitive NeuroscienceDocument6 pages1988 Perspectives On Cognitive NeuroscienceAna HortaNo ratings yet
- Consciousness in The Universe Is Scale Invariant and Implies An Event Horizon of The Human Brain PDFDocument39 pagesConsciousness in The Universe Is Scale Invariant and Implies An Event Horizon of The Human Brain PDFAdrián GonzálezNo ratings yet
- Science Abi8375Document5 pagesScience Abi8375Mateo MendozaNo ratings yet
- Forced Conceptual Thought Induced by Electrical Stimulation of The Left Prefrontal Gyrus Involves Widespread Neural NetworksDocument13 pagesForced Conceptual Thought Induced by Electrical Stimulation of The Left Prefrontal Gyrus Involves Widespread Neural NetworksGokudera HayatoNo ratings yet
- (1977) A Principle of Neural Assoiative MemoryDocument11 pages(1977) A Principle of Neural Assoiative MemorymartindsqNo ratings yet
- CPN 16 018Document14 pagesCPN 16 018Febria Rike ErlianaNo ratings yet
- From Sensation To Cognition: ReviewarticleDocument40 pagesFrom Sensation To Cognition: ReviewarticleAguelito AlbertNo ratings yet
- Pessoa NRN 2008 PDFDocument11 pagesPessoa NRN 2008 PDFatarNo ratings yet
- On The Relationship Between Emotion and CognitionDocument11 pagesOn The Relationship Between Emotion and CognitionLorenzo Maria CerviNo ratings yet
- Neurobiology of CognitionDocument4 pagesNeurobiology of CognitionNATARAJ REDDYNo ratings yet
- Perceptual Representations and The Vividness of Stimulus-Triggered and Stimulus-Independent Experiences (Fazenkas Et Al, 2021)Document26 pagesPerceptual Representations and The Vividness of Stimulus-Triggered and Stimulus-Independent Experiences (Fazenkas Et Al, 2021)GokushimakNo ratings yet
- Empathy CircuitsDocument9 pagesEmpathy Circuitscfrl25No ratings yet
- Review: The Reorienting System of The Human Brain: From Environment To Theory of MindDocument19 pagesReview: The Reorienting System of The Human Brain: From Environment To Theory of MindMarina Sánchez PicazoNo ratings yet
- 18 5 SteinmanDocument6 pages18 5 SteinmanPierre A. RodulfoNo ratings yet
- Gergely, G., & Csibra, G. (2006) - Sylvia's RecipeDocument15 pagesGergely, G., & Csibra, G. (2006) - Sylvia's RecipeVerónica BenavidezNo ratings yet
- Ferrari Et Al. (2012) Evidence For A Mirror Mechanism in Newborn MonkeysDocument14 pagesFerrari Et Al. (2012) Evidence For A Mirror Mechanism in Newborn MonkeysVerónica BenavidezNo ratings yet
- Short Communication Action Observation Activates Premotor and Parietal Areas in A Somatotopic Manner: An fMRI StudyDocument5 pagesShort Communication Action Observation Activates Premotor and Parietal Areas in A Somatotopic Manner: An fMRI StudyMARIA FERNANDA SANCHEZ HERNANDEZNo ratings yet
- Brain Network and Graph Theory PDFDocument13 pagesBrain Network and Graph Theory PDFVedant PatnaikNo ratings yet
- Neuroanatomy For The Psychiatrist: Textbook of PsychiatryDocument37 pagesNeuroanatomy For The Psychiatrist: Textbook of Psychiatrykrysdana22No ratings yet
- Advanced Kinesiology FinalDocument97 pagesAdvanced Kinesiology FinalTnum100% (1)
- Book Review - Electrodermal Responses What Happens in The BrainDocument12 pagesBook Review - Electrodermal Responses What Happens in The BrainamishcarNo ratings yet
- HypothalamusDocument39 pagesHypothalamusAfsaneh Javanmard100% (1)
- Types of NeuronsDocument10 pagesTypes of NeuronsJaspherBalderaNo ratings yet
- New Targets For RTMS in Depression - Review of Convergent Evidence - 2013Document10 pagesNew Targets For RTMS in Depression - Review of Convergent Evidence - 2013Lina SantosNo ratings yet
- Bullet To The BrainDocument4 pagesBullet To The BrainSmokethetree100% (4)
- Introduction To Physiology of The Nervous System (CLS, Allied Medical Student)Document34 pagesIntroduction To Physiology of The Nervous System (CLS, Allied Medical Student)Cristina PopescuNo ratings yet
- Katherine A. Sloman, Sigal Balshine, Rod W. Wilson Behaviour and Physiology of Fish Fish Physiology Volume 24 2005Document490 pagesKatherine A. Sloman, Sigal Balshine, Rod W. Wilson Behaviour and Physiology of Fish Fish Physiology Volume 24 2005Beatrice SavaNo ratings yet
- Human Development Across The Lifespan: Chapter 5 (2008)Document36 pagesHuman Development Across The Lifespan: Chapter 5 (2008)Jordan100% (3)
- Neuroanatómia Vizsgatételek A És B Csoport Képletekkel HZ ENDocument25 pagesNeuroanatómia Vizsgatételek A És B Csoport Képletekkel HZ ENTowan NguyenNo ratings yet
- Brain WebquestDocument3 pagesBrain WebquestCarlie100% (1)
- Nervous System Grade 6 ReviewerDocument12 pagesNervous System Grade 6 ReviewerGee ZNo ratings yet
- Memory SkillsDocument70 pagesMemory SkillsAsiri Samanatha Elegoda100% (1)
- 2.1.9 Practice Brain BiologyDocument3 pages2.1.9 Practice Brain Biologybuiclara07No ratings yet
- Güntürkün, Bugnyar (2016) Cognition Without CortexDocument13 pagesGüntürkün, Bugnyar (2016) Cognition Without CortexMak PavičevićNo ratings yet
- Neuroimaging - Cognitive and Clinical NeuroscienceDocument478 pagesNeuroimaging - Cognitive and Clinical NeuroscienceJosé Ramírez100% (3)
- A1. Functional Areas of Cerebral CortexDocument3 pagesA1. Functional Areas of Cerebral Cortexspindr07No ratings yet
- The Nervous SystemDocument54 pagesThe Nervous Systemeleldev100% (2)
- NervousDocument25 pagesNervousfairyzzzNo ratings yet
- Caring For The Patient With Cerebrovascular Disease Super FinalDocument84 pagesCaring For The Patient With Cerebrovascular Disease Super FinalDexter Niel Ortilano CPAC-SNNo ratings yet
- Individual Project - PresentationDocument55 pagesIndividual Project - PresentationTan Kar YeowNo ratings yet
- CVA Case StudyDocument20 pagesCVA Case Studybetchai18100% (5)
- Cerebral Cortex 1Document37 pagesCerebral Cortex 1Dorin PathakNo ratings yet
- Fisiología de La AtenciónDocument7 pagesFisiología de La AtenciónMaferez AC0% (1)
- Nervous SystemDocument94 pagesNervous Systemmr pcNo ratings yet
- RadionicsDocument178 pagesRadionicsNisha Joshi82% (22)
- AphasiaDocument29 pagesAphasiaHannan FarooqNo ratings yet
- The Olfactory PathwaysDocument3 pagesThe Olfactory PathwaysLeo TolentinoNo ratings yet
- Brain Bee R1 P2Document9 pagesBrain Bee R1 P2sarahsyedazakiNo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (28)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (81)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)No ratings yet
- The Marshmallow Test: Mastering Self-ControlFrom EverandThe Marshmallow Test: Mastering Self-ControlRating: 4.5 out of 5 stars4.5/5 (58)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (170)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- Self-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!From EverandSelf-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!Rating: 5 out of 5 stars5/5 (1)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingFrom EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingRating: 4 out of 5 stars4/5 (1138)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Summary: Hidden Potential: The Science of Achieving Greater Things By Adam Grant: Key Takeaways, Summary and AnalysisFrom EverandSummary: Hidden Potential: The Science of Achieving Greater Things By Adam Grant: Key Takeaways, Summary and AnalysisRating: 4.5 out of 5 stars4.5/5 (15)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Troubled: A Memoir of Foster Care, Family, and Social ClassFrom EverandTroubled: A Memoir of Foster Care, Family, and Social ClassRating: 4.5 out of 5 stars4.5/5 (27)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- The Secret of the Golden Flower: A Chinese Book Of LifeFrom EverandThe Secret of the Golden Flower: A Chinese Book Of LifeRating: 5 out of 5 stars5/5 (4)
- Summary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (61)