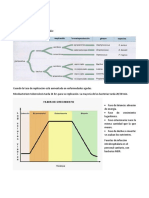
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Exploring The History of Neuropsychology-Selected Papers - Benton (2000)Document365 pagesExploring The History of Neuropsychology-Selected Papers - Benton (2000)AnadelRosarioNo ratings yet
- Apuntes de HematologíaDocument80 pagesApuntes de HematologíaJosé Manuel López RamírezNo ratings yet
- 00d03cb9bd283eb4 1Document87 pages00d03cb9bd283eb4 1José Manuel López RamírezNo ratings yet
- Esguince TobilloDocument39 pagesEsguince TobilloJosé Manuel López RamírezNo ratings yet
- Microbiota en La Salud y Enfermedad - En.esDocument11 pagesMicrobiota en La Salud y Enfermedad - En.esJosé Manuel López RamírezNo ratings yet
- CODODocument54 pagesCODOJosé Manuel López Ramírez100% (1)
- Nuevos Fármacos Antiepilépticos (II) : Avances en TerapéuticaDocument5 pagesNuevos Fármacos Antiepilépticos (II) : Avances en TerapéuticaJosé Manuel López RamírezNo ratings yet
- News Analysis: Denmark Says It'S Permanently Stopping Use of The Astrazeneca VaccineDocument23 pagesNews Analysis: Denmark Says It'S Permanently Stopping Use of The Astrazeneca VaccineJana HudhudNo ratings yet
- Human Blood CellsDocument3 pagesHuman Blood CellscsamarinaNo ratings yet
- Masaje TailandesDocument89 pagesMasaje TailandesEnderNo ratings yet
- Chapter 29Document93 pagesChapter 29b_comic9276No ratings yet
- Selecciones Reader S Digest Marzo 2012Document67 pagesSelecciones Reader S Digest Marzo 2012kevineli67% (3)
- WosaDocument18 pagesWosaAnusheelaNo ratings yet
- Protocolo Transplante de Higado Adultos Enero 2011Document9 pagesProtocolo Transplante de Higado Adultos Enero 2011alcontado_80No ratings yet
- 1.4 Butanediol AKA 1.4 BDODocument31 pages1.4 Butanediol AKA 1.4 BDOJohn PerryNo ratings yet
- AnemiaDocument9 pagesAnemiaMila Canoza HerreraNo ratings yet
- Protocolo Coledocolitiasis 1208258857789208 9Document11 pagesProtocolo Coledocolitiasis 1208258857789208 9David ZapataNo ratings yet
- Tongue in Complete DentureDocument8 pagesTongue in Complete DentureAman SachdevaNo ratings yet
- Tecnica Quirurgica Capitulo 1Document4 pagesTecnica Quirurgica Capitulo 1CAROL EIMI ORELLANA HURTADONo ratings yet
- Hospitals Patient Care Units and Ward ManagementDocument22 pagesHospitals Patient Care Units and Ward Managementbemina ja100% (1)
- Mine Rescue Questions and Answers PDFDocument19 pagesMine Rescue Questions and Answers PDFInayat ullah BalochNo ratings yet
- KeywordsDocument6,231 pagesKeywordshszforu0% (1)
- Reglamento para Establecimiento SaunaDocument7 pagesReglamento para Establecimiento SaunaAdolff PasacaNo ratings yet
- Traditional CHinese Medicine..Document30 pagesTraditional CHinese Medicine..Erlinda PosadasNo ratings yet
- No Tome Itraconazol NormonDocument8 pagesNo Tome Itraconazol NormonJosué EstradaNo ratings yet
- A Case Report - Surgical Relief of Colic in An EquineDocument4 pagesA Case Report - Surgical Relief of Colic in An Equinelinda nugrohoNo ratings yet
- Estructuras Por Alfredo EidelszteinDocument6 pagesEstructuras Por Alfredo EidelszteinMariaSoledadDiaz75% (4)
- Clave AmarillaDocument30 pagesClave AmarillaDajhanna Palacios NoriegaNo ratings yet
- Como É Praticada A Medicina Ayurveda Na ÍndiaDocument24 pagesComo É Praticada A Medicina Ayurveda Na ÍndiacmpmNo ratings yet
- Hubungan Beban Kerja Perawat Perawat Dengan Pelaksanaan Discharge Planning Diruang Rawat Inap Rumah Sakit Bhayangkara Palembang Tahun 2018Document10 pagesHubungan Beban Kerja Perawat Perawat Dengan Pelaksanaan Discharge Planning Diruang Rawat Inap Rumah Sakit Bhayangkara Palembang Tahun 2018Dedes SahpitraNo ratings yet
- SIG-PR-07 Ataques Ofidicos, Abejas Africanizadas y AvispasDocument7 pagesSIG-PR-07 Ataques Ofidicos, Abejas Africanizadas y AvispasMax GarffeNo ratings yet
- DEPRESIÓNDocument6 pagesDEPRESIÓNAriel Gonzalez LeonNo ratings yet
- Brochure Maquet Star IIDocument20 pagesBrochure Maquet Star IImikepower007No ratings yet
- Fer (SFBC)Document2 pagesFer (SFBC)DahmaniNo ratings yet
- Curso - Actualizacion Materno Fetal Clinic 2Document269 pagesCurso - Actualizacion Materno Fetal Clinic 2Rosália CoutadaNo ratings yet
- Funciones de La PielDocument3 pagesFunciones de La Pieldenisse AguasNo ratings yet