You might also like
- Classical Approach to Constrained and Unconstrained Molecular DynamicsFrom EverandClassical Approach to Constrained and Unconstrained Molecular DynamicsNo ratings yet
- 01 PDFDocument12 pages01 PDFMihai MihaiNo ratings yet
- Cgtase ThesisDocument7 pagesCgtase Thesisrhondacolemansavannah100% (2)
- Analyzing Heat Capacity Profiles of Peptide-Containing Membranes: Cluster Formation of Gramicidin ADocument13 pagesAnalyzing Heat Capacity Profiles of Peptide-Containing Membranes: Cluster Formation of Gramicidin Anurul9535No ratings yet
- Retention TImesDocument9 pagesRetention TImesNateNo ratings yet
- Characterizing Moisture Uptake and Plasticization Effects of Water On Amorphous Amylose Starch Models Using Molecular Dynamics MethodsDocument24 pagesCharacterizing Moisture Uptake and Plasticization Effects of Water On Amorphous Amylose Starch Models Using Molecular Dynamics MethodsIoNo ratings yet
- Application of Chemical Equilibrium Calculations For The Prediction of Ash-Induced AgglomerationDocument12 pagesApplication of Chemical Equilibrium Calculations For The Prediction of Ash-Induced AgglomerationteeeeeeeeeeeeeeeeeeeeeeeeeeeeettNo ratings yet
- On The Truncation of Long-Range Electrostatic Interactions in DNADocument17 pagesOn The Truncation of Long-Range Electrostatic Interactions in DNADavidFurmanNo ratings yet
- Energy Landscape of The Prion Protein Helix 1 Probed by Metadynamics and NMRDocument10 pagesEnergy Landscape of The Prion Protein Helix 1 Probed by Metadynamics and NMRVenkata Suryanarayana GorleNo ratings yet
- Molecular Recognition of Receptor Sites Using A PDFDocument11 pagesMolecular Recognition of Receptor Sites Using A PDFChely Romero LópezNo ratings yet
- Alexandrov Et Al. - 2008 - Microscale Fluorescent Thermal Stability Assay For Membrane ProteinsDocument9 pagesAlexandrov Et Al. - 2008 - Microscale Fluorescent Thermal Stability Assay For Membrane ProteinsErikNo ratings yet
- Chemical Engineering ScienceDocument7 pagesChemical Engineering ScienceclaudiosldfNo ratings yet
- Tratamiento de DatosDocument6 pagesTratamiento de DatosXaulo Del Rosario LecarosNo ratings yet
- Mass Transfer of Proteins in 2019Document6 pagesMass Transfer of Proteins in 2019NataliaNo ratings yet
- 2 Cross-Linking With Microbial Transglutaminase Relationship Between ... (5p)Document5 pages2 Cross-Linking With Microbial Transglutaminase Relationship Between ... (5p)Linh PhamNo ratings yet
- GPCR QsarDocument12 pagesGPCR QsarEdgarZuluagaNo ratings yet
- Use of Periodic Boundary Conditions To Calculate Accurate - Sheet Frequencies Using Density Functional TheoryDocument11 pagesUse of Periodic Boundary Conditions To Calculate Accurate - Sheet Frequencies Using Density Functional TheorybbteinsteinNo ratings yet
- Accurate Proton Affinity and Gas Phase Basicity Value For Molecular Important in BiocatalysisDocument11 pagesAccurate Proton Affinity and Gas Phase Basicity Value For Molecular Important in BiocatalysisNguyen NghiNo ratings yet
- Protein Unfolding Behavior Studied by Elastic Network ModelDocument11 pagesProtein Unfolding Behavior Studied by Elastic Network ModeljmxnetdjNo ratings yet
- Medium Optimization For Glutathione Production by 6isiae: Saccharomyces CereDocument7 pagesMedium Optimization For Glutathione Production by 6isiae: Saccharomyces CereAndréia AnschauNo ratings yet
- Molecular Modeling of The Enantioselectivity in Lipase-Catalyzed Transesterification ReactionsDocument12 pagesMolecular Modeling of The Enantioselectivity in Lipase-Catalyzed Transesterification ReactionsDavid AlvarezNo ratings yet
- A Simple Method For Displaying The Hydropathic Character of A ProteinDocument28 pagesA Simple Method For Displaying The Hydropathic Character of A ProteinBiosynthesisNo ratings yet
- Multivariate Curve Resolution: A Possible Tool in The Detection of Intermediate Structures in Protein FoldingDocument13 pagesMultivariate Curve Resolution: A Possible Tool in The Detection of Intermediate Structures in Protein Foldingakshdeep singhNo ratings yet
- A Mutation Data Matrix For Transmembrane Proteins: LettersDocument7 pagesA Mutation Data Matrix For Transmembrane Proteins: Letterscarlos ArozamenaNo ratings yet
- Exer 10Document7 pagesExer 10Yana CatherineNo ratings yet
- Acs Energyfuels 7b02507Document11 pagesAcs Energyfuels 7b02507Leonardo JaimesNo ratings yet
- 1 s2.0 S0006291X02002577 MainDocument6 pages1 s2.0 S0006291X02002577 MainRishab PandaNo ratings yet
- Metanol HeptanoDocument22 pagesMetanol HeptanoMary De la CruzNo ratings yet
- Poulet 2003Document7 pagesPoulet 2003Fatin Farhan HaqueNo ratings yet
- GIAO/DFT Evaluation of C NMR Chemical Shifts of Selected Acetals Based On DFT Optimized GeometriesDocument8 pagesGIAO/DFT Evaluation of C NMR Chemical Shifts of Selected Acetals Based On DFT Optimized Geometriesganesh_orcrdNo ratings yet
- Modelling Protein Docking Using Doking ImpDocument15 pagesModelling Protein Docking Using Doking Imptaoufik akabliNo ratings yet
- Alteration of Enzyme Specificity by Computational Loop Remodeling and DesignDocument6 pagesAlteration of Enzyme Specificity by Computational Loop Remodeling and DesignsuryasanNo ratings yet
- Protein-Polysaccharide InteractionsDocument13 pagesProtein-Polysaccharide Interactionsakking0905No ratings yet
- MJ ArticleDocument22 pagesMJ ArticlecptudorNo ratings yet
- A. 1992. Makhatadze, G. I. Protein Interactions With Urea and Guanidium ChlorideDocument15 pagesA. 1992. Makhatadze, G. I. Protein Interactions With Urea and Guanidium ChlorideJorge Luis RomeroNo ratings yet
- Review Gags MS 2 2009Document19 pagesReview Gags MS 2 2009Ariadna BiroccoNo ratings yet
- On The Accuracy of Homology Modeling and Sequence AlignmentDocument10 pagesOn The Accuracy of Homology Modeling and Sequence AlignmentCRISTIAN GABRIEL ZAMBRANO VEGANo ratings yet
- Hill 2004Document16 pagesHill 2004brouuorbNo ratings yet
- 10 1016@j Molliq 2019 111008Document8 pages10 1016@j Molliq 2019 111008muh. almusyafirNo ratings yet
- Determination of Lumped Rate Coefficients of Proteins: Effect of AdsorptionDocument6 pagesDetermination of Lumped Rate Coefficients of Proteins: Effect of AdsorptionCelso Hissao MaedaNo ratings yet
- A DFT-Based Computational Study On Hydrogen Tunneling Phenomenon in The Isomerization of MethylhydroxycarbeneDocument10 pagesA DFT-Based Computational Study On Hydrogen Tunneling Phenomenon in The Isomerization of MethylhydroxycarbenejumaidilawalNo ratings yet
- Vishnakov DPD ProteinsDocument7 pagesVishnakov DPD Proteinssoumava palitNo ratings yet
- Spectroscopic Study of Ca2+ Membrane-Induced Secondary Structural in Bovine Prothrombin FragmentDocument11 pagesSpectroscopic Study of Ca2+ Membrane-Induced Secondary Structural in Bovine Prothrombin Fragmentbelqis ratuNo ratings yet
- Mirzadeh - 2015 - Enhanced Protein Production in Escherichia Coli by Optimization of Cloning Scars at The Vector-Coding Sequence JunctionDocument7 pagesMirzadeh - 2015 - Enhanced Protein Production in Escherichia Coli by Optimization of Cloning Scars at The Vector-Coding Sequence JunctionmlNo ratings yet
- Full Length Article: SciencedirectDocument15 pagesFull Length Article: SciencedirectLuiza L. Dos AnjosNo ratings yet
- Ab Initio Characterization of Building UDocument67 pagesAb Initio Characterization of Building UIolanta BalanNo ratings yet
- Thermal Decomposition of Wheat, Oat, Barley, and Brassica: Carinata Straws. A Kinetic StudyDocument7 pagesThermal Decomposition of Wheat, Oat, Barley, and Brassica: Carinata Straws. A Kinetic StudyJoluis CcanahuireNo ratings yet
- Molecular Structure, Nbo and Homo-Lumo Analysis of Quercetin On Single Layer Graphene by Density Functional TheoryDocument9 pagesMolecular Structure, Nbo and Homo-Lumo Analysis of Quercetin On Single Layer Graphene by Density Functional TheoryAnonymous oSuBJMNo ratings yet
- 2017 Modeling of Biomass Productivity in Dense Microalgal Culture Using Computational Fluid DynamicsDocument8 pages2017 Modeling of Biomass Productivity in Dense Microalgal Culture Using Computational Fluid Dynamicsmorillas.ainoNo ratings yet
- Mechanisms of Protein FoldingDocument14 pagesMechanisms of Protein Foldingzaid azharNo ratings yet
- Molecular Dynamics On Laccase From Trametes Versicolor To Examine Thermal Stabibity Induced by Salt BridgesDocument12 pagesMolecular Dynamics On Laccase From Trametes Versicolor To Examine Thermal Stabibity Induced by Salt BridgesArturo RojoNo ratings yet
- Ye jcb07Document15 pagesYe jcb07suryasanNo ratings yet
- Proton Transfer Reactions in Native and Deionized Bacteriorhodopsin Upon Delipidation and MonomerizationDocument9 pagesProton Transfer Reactions in Native and Deionized Bacteriorhodopsin Upon Delipidation and MonomerizationJasa ProwebNo ratings yet
- Asboth 2000Document18 pagesAsboth 2000Nguyễn Đức TùngNo ratings yet
- 2017.09.11 - VARGAS - ROY - CTC - Study of The Stability of Small Urh Clusters Found by A Genetic AlgorithmDocument8 pages2017.09.11 - VARGAS - ROY - CTC - Study of The Stability of Small Urh Clusters Found by A Genetic AlgorithmAlejandra AwimbaweNo ratings yet
- Journal of Saudi Chemical Society: King Saud UniversityDocument7 pagesJournal of Saudi Chemical Society: King Saud UniversityLinda SetyawatiNo ratings yet
- Aspergillus Terreus Through Introduction of Disulfide BondsDocument29 pagesAspergillus Terreus Through Introduction of Disulfide BondsSHREYANo ratings yet
- Waveform Inversion of Amplitude-Versus-Offset Data Using A Genetic Algorithm SL2.7Document4 pagesWaveform Inversion of Amplitude-Versus-Offset Data Using A Genetic Algorithm SL2.7a1341365No ratings yet
- Byler 1988Document16 pagesByler 1988Amuthachelvi DanielNo ratings yet
- Three-Dimensional Structure of P-GlycoproteinDocument6 pagesThree-Dimensional Structure of P-GlycoproteinSakshi IssarNo ratings yet
- CDC 105136 DS1Document77 pagesCDC 105136 DS1Leidy Constanza Villalobos GonzalezNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFLeidy Constanza Villalobos GonzalezNo ratings yet
- Paxton 2019Document18 pagesPaxton 2019Leidy Constanza Villalobos GonzalezNo ratings yet
- Rodbard 1971Document40 pagesRodbard 1971Leidy Constanza Villalobos GonzalezNo ratings yet
- HHS Public Access: Quantitation of NAD Biosynthesis From The Salvage Pathway inDocument12 pagesHHS Public Access: Quantitation of NAD Biosynthesis From The Salvage Pathway inLeidy Constanza Villalobos GonzalezNo ratings yet
- 1 s2.0 S0021925819316795 MainDocument7 pages1 s2.0 S0021925819316795 MainLeidy Constanza Villalobos GonzalezNo ratings yet
- 10 1039@c9an02553gDocument10 pages10 1039@c9an02553gLeidy Constanza Villalobos GonzalezNo ratings yet
- Location, Location, Location: Compartmentalization of NAD Synthesis and Functions in Mammalian CellsDocument16 pagesLocation, Location, Location: Compartmentalization of NAD Synthesis and Functions in Mammalian CellsLeidy Constanza Villalobos GonzalezNo ratings yet
- GKP 854Document14 pagesGKP 854Leidy Constanza Villalobos GonzalezNo ratings yet
- Magni2004 Article EnzymologyOfNADHomeostasisInMaDocument16 pagesMagni2004 Article EnzymologyOfNADHomeostasisInMaLeidy Constanza Villalobos GonzalezNo ratings yet
- JVC HR-S6970AA Service ManualDocument61 pagesJVC HR-S6970AA Service ManualPriyantha GanegodaNo ratings yet
- Marketing Case StudiesDocument10 pagesMarketing Case StudiesSwati ChamariaNo ratings yet
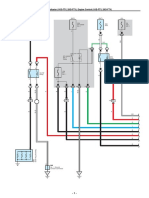
- ECT and A/T Indicator (1KD-FTV, 2KD-FTV), Engine Control (1KD-FTV, 2KD-FTV)Document33 pagesECT and A/T Indicator (1KD-FTV, 2KD-FTV), Engine Control (1KD-FTV, 2KD-FTV)Erick Lizana Neyra100% (4)
- Pili Danica - Dyners Corporation - Case Analysis #1Document2 pagesPili Danica - Dyners Corporation - Case Analysis #1DANICA DANIELA PILINo ratings yet
- Garden Namkeen TestDocument12 pagesGarden Namkeen Testramakrishna.goliNo ratings yet
- 12th Commerce Study Material English MediumDocument97 pages12th Commerce Study Material English MediumDeepak ajar100% (1)
- Prs-021-0025 - C. Melchers - Coca Cola Naga ProjectDocument2 pagesPrs-021-0025 - C. Melchers - Coca Cola Naga ProjectMiko Christopher OroyanNo ratings yet
- Export Value DeclarationDocument1 pageExport Value DeclarationdocrmpjodNo ratings yet
- Mouthwash, PEG, Sulfate, Betaine FreeDocument2 pagesMouthwash, PEG, Sulfate, Betaine FreerekhilaNo ratings yet
- Final CV-Europass-20190916-MezakMatijević-EN (2) - Kopija PDFDocument4 pagesFinal CV-Europass-20190916-MezakMatijević-EN (2) - Kopija PDFMirela Mezak StastnyNo ratings yet
- English Grammar ExerciseDocument4 pagesEnglish Grammar Exerciseanca_2402No ratings yet
- MOI Lesson PlanDocument4 pagesMOI Lesson PlanPreeti KumariNo ratings yet
- Master Teacher Developmental Plan Real2021 2022Document6 pagesMaster Teacher Developmental Plan Real2021 2022Roygvib Clemente MontañoNo ratings yet
- I Am A SEAL Team Six Warrior Memoirs of An American SoldierDocument12 pagesI Am A SEAL Team Six Warrior Memoirs of An American SoldierMacmillan Publishers43% (51)
- GaugesDocument33 pagesGaugesUsman ansarNo ratings yet
- Geotecnical Limit State AustralianDocument6 pagesGeotecnical Limit State Australianprashanth c.nNo ratings yet
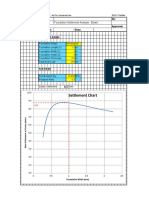
- Settlement Chart: Foundation Settlement Analysis - ElasticDocument2 pagesSettlement Chart: Foundation Settlement Analysis - ElasticCSEC Uganda Ltd.No ratings yet
- Decision Tree PrimerDocument42 pagesDecision Tree PrimerMuzzamil JanjuaNo ratings yet
- August 2009 Go Guide Newsletter The MountaineersDocument20 pagesAugust 2009 Go Guide Newsletter The MountaineersMountaineersNo ratings yet
- Language Assessment in Year 2 KSSRDocument13 pagesLanguage Assessment in Year 2 KSSRCahaya Hati100% (1)
- 29102015000000B - Boehler DCMS-IG - SWDocument1 page29102015000000B - Boehler DCMS-IG - SWErdinc BayatNo ratings yet
- Theoretical Foundation For Large-Scale Computations For Nonlinear Material BehaviorDocument411 pagesTheoretical Foundation For Large-Scale Computations For Nonlinear Material BehaviorDalelAzaiezNo ratings yet
- UntitledDocument42 pagesUntitledDiana JacquelineNo ratings yet
- The Philippine Informal Reading InventoryDocument2 pagesThe Philippine Informal Reading InventoryVenilyn Cruz91% (11)
- Simulación de Eventos Discretos Eduardo García Dunna May 4, 2015Document22 pagesSimulación de Eventos Discretos Eduardo García Dunna May 4, 2015Andrea GutierrezNo ratings yet
- Readings in Philippine History-CeDocument11 pagesReadings in Philippine History-Cedeluna.jerremieNo ratings yet
- D.ANDAN PS-PROJECT WATCH ActionPlan 2018-2019Document3 pagesD.ANDAN PS-PROJECT WATCH ActionPlan 2018-2019Maria Fe PanuganNo ratings yet
- Adult Neurological QuestionnaireDocument1 pageAdult Neurological QuestionnaireIqbal BaryarNo ratings yet
- Production Technology of Cow PeaDocument20 pagesProduction Technology of Cow PeaMUHAMMAD AQEELNo ratings yet
- Quantity Calculations - LPG and Chemical GasesDocument35 pagesQuantity Calculations - LPG and Chemical GasesOsama SalamaNo ratings yet