You might also like
- Carbon Capture Technologies for Gas-Turbine-Based Power PlantsFrom EverandCarbon Capture Technologies for Gas-Turbine-Based Power PlantsNo ratings yet
- 1 s2.0 S0196890421006671 MainDocument10 pages1 s2.0 S0196890421006671 MainWei WangNo ratings yet
- 2019 Kreitz - Dynamic Simulation of The CO2 Methanation in A Micro-Structured Fixed-Bed ReactorDocument51 pages2019 Kreitz - Dynamic Simulation of The CO2 Methanation in A Micro-Structured Fixed-Bed Reactoranilaltindas093No ratings yet
- Energy Conversion and ManagementDocument9 pagesEnergy Conversion and ManagementAbeer El ShahawyNo ratings yet
- Wierzbicki Et Al. (2014) - Combustion of Propane With PT and RH Catalysts in A Meso-Scale Heat Recirculating CombuDocument7 pagesWierzbicki Et Al. (2014) - Combustion of Propane With PT and RH Catalysts in A Meso-Scale Heat Recirculating CombuEmily LinNo ratings yet
- Performance Analysis of Operational Strategies For Mono - 2020 - International JDocument13 pagesPerformance Analysis of Operational Strategies For Mono - 2020 - International JDuvan Leandro Parra VelandiaNo ratings yet
- 1 s2.0 S0360319923062432 MainDocument9 pages1 s2.0 S0360319923062432 MainFlames ShoppingNo ratings yet
- Analisis de Metil-Tol para La Produccion de Combustible para Las Centrales ElectricasDocument9 pagesAnalisis de Metil-Tol para La Produccion de Combustible para Las Centrales ElectricasEDUAR SEBASTIAN BUENO SEGURANo ratings yet
- Simulación Numérica de La Pirólisis Rápida de Biomasa en Un Reactor SinfínDocument9 pagesSimulación Numérica de La Pirólisis Rápida de Biomasa en Un Reactor SinfínJasivis PoloNo ratings yet
- Conceptual Design and Analysis of A Novel Process For Hydrogen Liquefaction PDFDocument24 pagesConceptual Design and Analysis of A Novel Process For Hydrogen Liquefaction PDFDilla WahabNo ratings yet
- Duarte Pereira de Araújo Coutinho ThermoeconomicDocument10 pagesDuarte Pereira de Araújo Coutinho ThermoeconomicJonathan ManzakiNo ratings yet
- HTPRDocument11 pagesHTPRDiego MaportiNo ratings yet
- Akeem K. Olaleye, Kunle J. Adedayo, Chunfei Wu, Mohamad A. Nahil, Meihong Wang, Paul T. WilliamsDocument11 pagesAkeem K. Olaleye, Kunle J. Adedayo, Chunfei Wu, Mohamad A. Nahil, Meihong Wang, Paul T. WilliamsVashisht SewsaransingNo ratings yet
- Performance Improvement of Pulverized Coal Fired Thermal Power Plant: A Retrofitting OptionDocument9 pagesPerformance Improvement of Pulverized Coal Fired Thermal Power Plant: A Retrofitting OptionOnatNo ratings yet
- HP4a 13 Giaconia Rev0607Document7 pagesHP4a 13 Giaconia Rev0607kkakosimNo ratings yet
- Topsoe Biogas ReformingDocument9 pagesTopsoe Biogas ReformingDiego MaportiNo ratings yet
- Manuscript - Part 1 - CompositeDocument36 pagesManuscript - Part 1 - CompositeMRNo ratings yet
- Bioenergy With Carbon Emissions Capture and Utilisation Towards GHG Neutrality PowertoGas Storage Via Hydrothermal Gasification2020applied EnergyDocument16 pagesBioenergy With Carbon Emissions Capture and Utilisation Towards GHG Neutrality PowertoGas Storage Via Hydrothermal Gasification2020applied EnergyAlejandro Andres RomeroNo ratings yet
- Water 15 03039 With CoverDocument21 pagesWater 15 03039 With Covernuli chanNo ratings yet
- Mathematical Modelling of A Hydrocracking Reactor For Triglyceride Conversion To Biofuel: Model Establishment and ValidationDocument11 pagesMathematical Modelling of A Hydrocracking Reactor For Triglyceride Conversion To Biofuel: Model Establishment and ValidationMarcelo Medrano TapiaNo ratings yet
- Solar and Biomass Hybridization Through Hydrothermal CarbonizationDocument12 pagesSolar and Biomass Hybridization Through Hydrothermal CarbonizationHarry Laksono NugrohoNo ratings yet
- Levebre 2019Document9 pagesLevebre 2019Ramadika AdrianNo ratings yet
- Thermodynamic Optimization of Bottoming CycleDocument15 pagesThermodynamic Optimization of Bottoming CycleStjepkoKatulićNo ratings yet
- Er-Rbib Bouallou Energy 2014Document9 pagesEr-Rbib Bouallou Energy 2014Yogendran GanesanNo ratings yet
- Synopsis: (PRN No. 201502677)Document12 pagesSynopsis: (PRN No. 201502677)sumitNo ratings yet
- Catalyst Regeneration Techniques inDocument8 pagesCatalyst Regeneration Techniques inCHE.ENG1734No ratings yet
- HRSG DesignDocument11 pagesHRSG DesignVennila RangithNo ratings yet
- Electrical Power and Energy Systems: A. Baghernejad, M. Yaghoubi, K. JafarpurDocument14 pagesElectrical Power and Energy Systems: A. Baghernejad, M. Yaghoubi, K. JafarpurAbderrahman HMAITINo ratings yet
- He (2016)Document13 pagesHe (2016)Suênia FernandesNo ratings yet
- Energy: Youfu Ma, Lijuan Yang, Junfu Lu, Yufeng PeiDocument12 pagesEnergy: Youfu Ma, Lijuan Yang, Junfu Lu, Yufeng PeiFajar Firstya AdamNo ratings yet
- Production of Gaseous Carriers Via Biomass GasificDocument10 pagesProduction of Gaseous Carriers Via Biomass GasificNicholas WilliamNo ratings yet
- Fixed Bed Journal 3Document11 pagesFixed Bed Journal 3Daniel EdohNo ratings yet
- Analysis of Temperature Glide Matching of Heat Pumps With Zeotropic Working Fluid Mixtures For Different Temperature GlidesDocument11 pagesAnalysis of Temperature Glide Matching of Heat Pumps With Zeotropic Working Fluid Mixtures For Different Temperature GlidesRodrigo Salgado PradenasNo ratings yet
- Paper 4Document9 pagesPaper 4Kátia Sofia GomesNo ratings yet
- Energy Conversion and Management: Huy Quoc Nguyen, Bahman ShabaniDocument29 pagesEnergy Conversion and Management: Huy Quoc Nguyen, Bahman ShabaniCarlos JavierNo ratings yet
- Exergetic Assessment of CO2 Methanation Processes For The Chemical StorageDocument12 pagesExergetic Assessment of CO2 Methanation Processes For The Chemical StorageAditya KarpeNo ratings yet
- 1 s2.0 S0196890420302016 MainDocument19 pages1 s2.0 S0196890420302016 MainMichael WangNo ratings yet
- 1 s2.0 S019689042100861X MainDocument15 pages1 s2.0 S019689042100861X MainxorrudslaNo ratings yet
- Carbon Capture Retrofit Options With The On-Site Addition of Gas Turbine Combined Heat and Power CycleDocument9 pagesCarbon Capture Retrofit Options With The On-Site Addition of Gas Turbine Combined Heat and Power CycleGustavo Ramirez AriasNo ratings yet
- CompressionDocument17 pagesCompressionSoroosh ZareNo ratings yet
- Exergy-Energy Analysis of Full Repowering of A Steam Power PlantDocument14 pagesExergy-Energy Analysis of Full Repowering of A Steam Power Plantsevero97No ratings yet
- J Enconman 2016 09 080Document11 pagesJ Enconman 2016 09 080RAHMAN MARUF MORSHED, 180011237No ratings yet
- Ijetae Icertsd 0213 90Document7 pagesIjetae Icertsd 0213 90rajNo ratings yet
- Literature Review of Differet Geothermal Fields Available in WorldDocument15 pagesLiterature Review of Differet Geothermal Fields Available in WorldVishnu Pratap Singh100% (1)
- Optimum Superheat Utilization of Extraction Steam in Double ReheatDocument10 pagesOptimum Superheat Utilization of Extraction Steam in Double ReheatAhmed AsaadNo ratings yet
- 3-Energy Efficiency Improvement For Industrial Boilers Through A Flue-Gas Condensing Heat Recovery System With Nonlinear MPC ApproachDocument15 pages3-Energy Efficiency Improvement For Industrial Boilers Through A Flue-Gas Condensing Heat Recovery System With Nonlinear MPC ApproachBerkefedeNo ratings yet
- Bioresource Technology: Manu Agarwal, James Tardio, S. Venkata MohanDocument6 pagesBioresource Technology: Manu Agarwal, James Tardio, S. Venkata MohanCarlos AlvarezNo ratings yet
- Exergy Economic and Life-Cycle Assessment of ORC SDocument23 pagesExergy Economic and Life-Cycle Assessment of ORC SChurio Silvera OscarNo ratings yet
- Prediction of Species Concentration in Syngas Produced Through Gasification of Different Bamboo Biomasses: A Numerical ApproachDocument12 pagesPrediction of Species Concentration in Syngas Produced Through Gasification of Different Bamboo Biomasses: A Numerical ApproachBELAJAR BERSAMA. NETNo ratings yet
- Full TextDocument12 pagesFull Textmahdi.shahparastiNo ratings yet
- Sdarticle 1Document8 pagesSdarticle 1Ravikiran TatavarthyNo ratings yet
- 1 s2.0 S0960148118315349 MainDocument13 pages1 s2.0 S0960148118315349 Mainrsiddharth2008No ratings yet
- Chemical Engineering Journal: Contents Lists Available atDocument10 pagesChemical Engineering Journal: Contents Lists Available atrafael lopezNo ratings yet
- Energy Conversion and ManagementDocument8 pagesEnergy Conversion and ManagementkyongngNo ratings yet
- Heat Transfer Studies at Different Speeds and Loads of Regenerative Air Preheater in Thermal Power PlantDocument6 pagesHeat Transfer Studies at Different Speeds and Loads of Regenerative Air Preheater in Thermal Power PlantdavidNo ratings yet
- 1 s2.0 S1385894713013302 MainDocument7 pages1 s2.0 S1385894713013302 MainAlex GutierrezNo ratings yet
- Fuel Processing Technology: Jonathan L. Wagner, Chien D. Le, Valeska P. Ting, Christopher J. ChuckDocument10 pagesFuel Processing Technology: Jonathan L. Wagner, Chien D. Le, Valeska P. Ting, Christopher J. ChuckJorgeWashiHuamanChjutalliNo ratings yet
- Renewable Energy: Mohamed Ali Masmoudi, Melik Sahraoui, Najla Grioui, Kamel HalouaniDocument11 pagesRenewable Energy: Mohamed Ali Masmoudi, Melik Sahraoui, Najla Grioui, Kamel HalouaniSaif EvonyNo ratings yet
- 1-S2.0-S0255270196041682-Main Westerterp PDFDocument11 pages1-S2.0-S0255270196041682-Main Westerterp PDFnicocarraraNo ratings yet
- Energy: Yusuf Bicer, Ibrahim DincerDocument10 pagesEnergy: Yusuf Bicer, Ibrahim DincerMuhammad OsamaNo ratings yet
- Development of Assessment Methods For Alternative Fuels For Naval Diesel EnginesDocument35 pagesDevelopment of Assessment Methods For Alternative Fuels For Naval Diesel Engineswilfred gomezNo ratings yet
- Phosgene Msds E4641ttDocument7 pagesPhosgene Msds E4641ttArif Adi NugrohoNo ratings yet
- Test Bank For Organic Chemistry 11th Edition Francis Carey Robert Giuliano Janice SmithDocument32 pagesTest Bank For Organic Chemistry 11th Edition Francis Carey Robert Giuliano Janice Smithxavianhatmgzmz9No ratings yet
- Artigo Sobre EstribosDocument5 pagesArtigo Sobre EstribosPaulo CotteNo ratings yet
- Biodegradable Plastic of Jicama Starch (Pachyrhizus Erosus) With Precipitate Calcium Carbonate As A FillerDocument5 pagesBiodegradable Plastic of Jicama Starch (Pachyrhizus Erosus) With Precipitate Calcium Carbonate As A FillerMonaliza MandacNo ratings yet
- Two Solids Lab PDFDocument1 pageTwo Solids Lab PDFLukeNo ratings yet
- GERODUR - HDPE Pipe and FittingsDocument107 pagesGERODUR - HDPE Pipe and FittingsJacky TamNo ratings yet
- SDS Nickel Cadmium PP ContainersDocument6 pagesSDS Nickel Cadmium PP ContainersharmlesdragonNo ratings yet
- The Nitrogen Cycle: of Microbes and MenDocument16 pagesThe Nitrogen Cycle: of Microbes and Mensohail1jadoonNo ratings yet
- Tablettng With Video-1Document40 pagesTablettng With Video-1Rahima Akter RakhiNo ratings yet
- 1 Laws Physcie 1STQTRDocument5 pages1 Laws Physcie 1STQTRKayzelle RefamonteNo ratings yet
- Intranasal Liposomes: An Approach For Drug Delivery To BrainDocument13 pagesIntranasal Liposomes: An Approach For Drug Delivery To BrainMaruf Momin100% (1)
- Gas Cylinder List WITH FITTING DETAILSDocument1 pageGas Cylinder List WITH FITTING DETAILSSameer SaeedNo ratings yet
- Derakane - Tech SupportDocument32 pagesDerakane - Tech SupportJainam ShahNo ratings yet
- 1-Handling of SolidsDocument10 pages1-Handling of SolidsAgus Wahyudhi100% (1)
- Electrostatic Ion Thrusters Xenon Ionized Cathode FilamentDocument2 pagesElectrostatic Ion Thrusters Xenon Ionized Cathode FilamentParama GuruNo ratings yet
- Unit III Humidity and SaturationDocument50 pagesUnit III Humidity and SaturationSaurav SatsangiNo ratings yet
- Topicwise Test Schedule of AIATS For AIPMT / AIIMS 2016: For Class 12th Studying / Passed StudentsDocument2 pagesTopicwise Test Schedule of AIATS For AIPMT / AIIMS 2016: For Class 12th Studying / Passed StudentsPranav GargNo ratings yet
- Lesson 1-2-3 Subject Methods ChemistryDocument13 pagesLesson 1-2-3 Subject Methods ChemistrykimaniNo ratings yet
- Transfluthrin WHODocument20 pagesTransfluthrin WHOYudhytha AnggarhaniNo ratings yet
- DT202B THERMAL TOP P 200 RH01 HONEY GLASSINE 65 UPM Raflatac SPADocument2 pagesDT202B THERMAL TOP P 200 RH01 HONEY GLASSINE 65 UPM Raflatac SPArcNo ratings yet
- Heat and Temperature NotesDocument2 pagesHeat and Temperature NotesMARISTELA MACARANASNo ratings yet
- MedPhysTerm-KSMP First Version (200906)Document216 pagesMedPhysTerm-KSMP First Version (200906)Ilkuk EomNo ratings yet
- Submitted by Submitted To: Adit Dubhashe MR - Rajesh JainDocument17 pagesSubmitted by Submitted To: Adit Dubhashe MR - Rajesh JainalpanaNo ratings yet
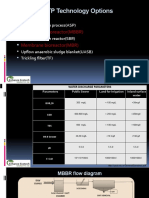
- STP Technology Options: Moving Bed Bioreactor (MBBR) Membrane Bioreactor (MBR)Document6 pagesSTP Technology Options: Moving Bed Bioreactor (MBBR) Membrane Bioreactor (MBR)amzad aliNo ratings yet
- 3m Petrifilm Rapid e Coli Coliform Count Plate Interpretation Guide RECDocument6 pages3m Petrifilm Rapid e Coli Coliform Count Plate Interpretation Guide RECLuz Katherine MartinezNo ratings yet
- Vortex Tube - Yunpeng XueDocument121 pagesVortex Tube - Yunpeng XueVladan MilojevićNo ratings yet
- prEN 10149-1 (2011) PDFDocument16 pagesprEN 10149-1 (2011) PDFneiva201950% (2)
- Compressible Fluid FlowDocument26 pagesCompressible Fluid FlowPradip GuptaNo ratings yet
- Lecture 8 Creep and RuptureDocument8 pagesLecture 8 Creep and RuptureAzhar AliNo ratings yet
- Practical Reservoir Engineering and CharacterizationFrom EverandPractical Reservoir Engineering and CharacterizationRating: 4.5 out of 5 stars4.5/5 (3)
- Well Control for Completions and InterventionsFrom EverandWell Control for Completions and InterventionsRating: 4 out of 5 stars4/5 (10)
- Pocket Guide to Flanges, Fittings, and Piping DataFrom EverandPocket Guide to Flanges, Fittings, and Piping DataRating: 3.5 out of 5 stars3.5/5 (22)
- Gas-Liquid And Liquid-Liquid SeparatorsFrom EverandGas-Liquid And Liquid-Liquid SeparatorsRating: 3.5 out of 5 stars3.5/5 (3)
- Machine Learning Guide for Oil and Gas Using Python: A Step-by-Step Breakdown with Data, Algorithms, Codes, and ApplicationsFrom EverandMachine Learning Guide for Oil and Gas Using Python: A Step-by-Step Breakdown with Data, Algorithms, Codes, and ApplicationsRating: 4 out of 5 stars4/5 (4)
- Machine Learning and Data Science in the Oil and Gas Industry: Best Practices, Tools, and Case StudiesFrom EverandMachine Learning and Data Science in the Oil and Gas Industry: Best Practices, Tools, and Case StudiesPatrick BangertRating: 3 out of 5 stars3/5 (2)
- Well Testing Project Management: Onshore and Offshore OperationsFrom EverandWell Testing Project Management: Onshore and Offshore OperationsNo ratings yet
- Biostratigraphic and Geological Significance of Planktonic ForaminiferaFrom EverandBiostratigraphic and Geological Significance of Planktonic ForaminiferaRating: 4 out of 5 stars4/5 (5)
- Advanced Biomass Gasification: New Concepts for Efficiency Increase and Product FlexibilityFrom EverandAdvanced Biomass Gasification: New Concepts for Efficiency Increase and Product FlexibilityRating: 3 out of 5 stars3/5 (2)
- Advanced Production Decline Analysis and ApplicationFrom EverandAdvanced Production Decline Analysis and ApplicationRating: 3.5 out of 5 stars3.5/5 (4)
- Asphaltene Deposition Control by Chemical Inhibitors: Theoretical and Practical ProspectsFrom EverandAsphaltene Deposition Control by Chemical Inhibitors: Theoretical and Practical ProspectsNo ratings yet
- Mooring System Engineering for Offshore StructuresFrom EverandMooring System Engineering for Offshore StructuresRating: 5 out of 5 stars5/5 (1)
- Essentials of Coating, Painting, and Lining for the Oil, Gas and Petrochemical IndustriesFrom EverandEssentials of Coating, Painting, and Lining for the Oil, Gas and Petrochemical IndustriesRating: 5 out of 5 stars5/5 (5)
- Heat Exchanger Equipment Field Manual: Common Operating Problems and Practical SolutionsFrom EverandHeat Exchanger Equipment Field Manual: Common Operating Problems and Practical SolutionsRating: 4 out of 5 stars4/5 (6)
- Gas and Oil Reliability Engineering: Modeling and AnalysisFrom EverandGas and Oil Reliability Engineering: Modeling and AnalysisRating: 4.5 out of 5 stars4.5/5 (6)
- A Practical Guide to Piping and Valves for the Oil and Gas IndustryFrom EverandA Practical Guide to Piping and Valves for the Oil and Gas IndustryRating: 5 out of 5 stars5/5 (3)
- Asset Integrity Management for Offshore and Onshore StructuresFrom EverandAsset Integrity Management for Offshore and Onshore StructuresNo ratings yet